大鼠颈动脉球囊损伤模型内膜增生过程中连接蛋白的变化
2020-05-21潘乐门陈帆风苏翔杨法镜
潘乐门 陈帆风 苏翔 杨法镜
血管内皮损伤可诱导动脉内膜增生,引起经皮动脉腔内治疗术后的动脉再狭窄。当血管内皮损伤时,血管平滑肌细胞可从中膜层向内膜层迁移,并增殖、分泌细胞外基质,形成再狭窄性病变[1]。经皮腔内治疗是针对动脉硬化狭窄疾病的重要治疗方法,而腔内治疗面临的主要问题在于如何保持术后的长期通畅率。据估计,近25%的动脉狭窄患者会在血管腔内术后1年内发生再狭窄。内膜增生导致的再狭窄问题,是影响动脉腔内治疗术后长期通畅率的重要因素[2]。因此,了解内膜增生过程的机制对于控制术后再狭窄,改善腔内手术效果非常重要。血管内皮细胞与血管平滑肌细胞之间的缝隙连接等细胞间通讯对血管壁的功能至关重要。在动脉粥样硬化的发展过程中,在血管内皮细胞与血管平滑肌细胞之间的接触点可以观察到连接蛋白(connexin,Cx)表达的逐渐变化,并且在动脉粥样硬化斑块中可以检测到连接蛋白表达模式的改变[3]。表明连接蛋白参与动脉粥样硬化的发展,这种分子是否参与球囊损伤引起的内膜增生和再狭窄的报道较少。在本研究中,通过球囊损伤后再狭窄模型来研究连接蛋白在内膜增生过程中的作用,并初步探索其相关机制。
1 资料与方法
1.1 模型建立 雄性SD大鼠(250±10)g购自北京生命之河实验动物有限公司,以标准的12h明暗循环进行饲养。将48只大鼠随机分为对照组(BI)和非损伤组(NI),每组各24只。对照组(BI)采用异氟醚麻醉大鼠,暴露左侧颈总动脉,用近端和远端动脉夹暂时阻断。将1.5mm球囊导管插入颈动脉并充气至6atm,然后在短距离内来回拉动3次,以产生内皮损伤。非损伤组(NI)以假手术的大鼠为对照。皮下注射止痛药和抗生素,恢复期常规监测大鼠。
1.2 组织病理学评价 术后用4%多聚甲醛固定颈动脉,石蜡包埋,沿损伤段行连续切片(2μm),苏木精-伊红(H&E)染色检测内膜增生,利用NIH图像处理系统获得内膜、中膜和管腔面积的定量。颈动脉切片进行增殖细胞核抗原(PCNA)染色。用微波加热提取抗原,4%正常山羊血清加1%牛血清白蛋白(BSA)在PBS中进行非特异性阻断30min,然后用抗大鼠PCNA兔多克隆抗体(1∶100)孵育切片。二抗选用二氨基联苯胺底物复合物酶标染色进行信号检测。最后,用苏木精对载玻片进行复染和分析。
1.3 Western blot 颈动脉组织加入电泳缓冲液匀化,再经电泳分离,转膜固定后,加入1∶1000稀释的Cx30、Cx37、Cx40、Cx43、ERK、Akt、(p)-ERK 和(p)-Akt的抗体在4℃孵育过夜,然后在室温下加入二抗中进一步孵育1h。用Image J软件计算磷酸化与总蛋白表达的比率。
1.4 数据处理 所有实验均用GraphPad Prism 6进行定量分析,结果以()表示,并采用t检验或单因素方差分析,随后进行Tukey后检验。P<0.05为差异具有统计学意义。
2 结果
2.1 连接蛋白的时间表达 在整个研究期间,BI和NI大鼠术后均保持健康,体重无明显下降。两组分别于术后6h、24h、7d、14d处死6只大鼠,进行Cx30、Cx37、Cx40和Cx43的检测。Western blot结果表明,随着时间的推移,Cx37表达呈下降趋势,Cx43表达呈上升趋势,而Cx30和Cx40表达无显著差异(见图1)。结果表明,在四种连接蛋白中,只有Cx37和Cx43在球囊损伤后的整个过程中表达有明显差异。
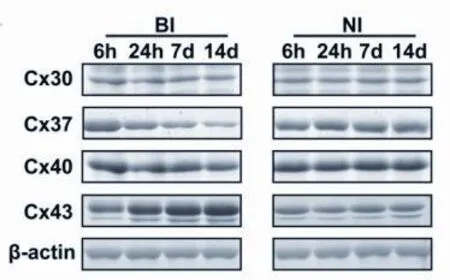
图1 连接蛋白的时间表达情况。为Cx30、Cx37、Cx40和Cx43这四种连接蛋白在术后6h、24h、7d和14d等4个时间点的Western印迹分析
2.2 球囊损伤致新生内膜形成 根据连接蛋白表达的结果,通过HE染色观察术后第14天球囊损伤对新生内膜形成的影响(每组6只)。在接受假手术的大鼠中未观察到明显的内膜增生,而在BI组中发现显著的内膜增生(见图2A)。定量形态计量学评估显示,BI组的内膜面积(见图2B,P<0.01)和内膜-中膜(I/M)比值(见图2E,P<0.01)较NI组显著增加,且BI组的管腔面积明显低于NI组(见图2D,P<0.01),而两组之间的内外膜间的区域无显著差异(见图2C)。
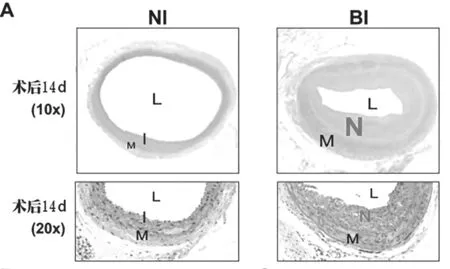
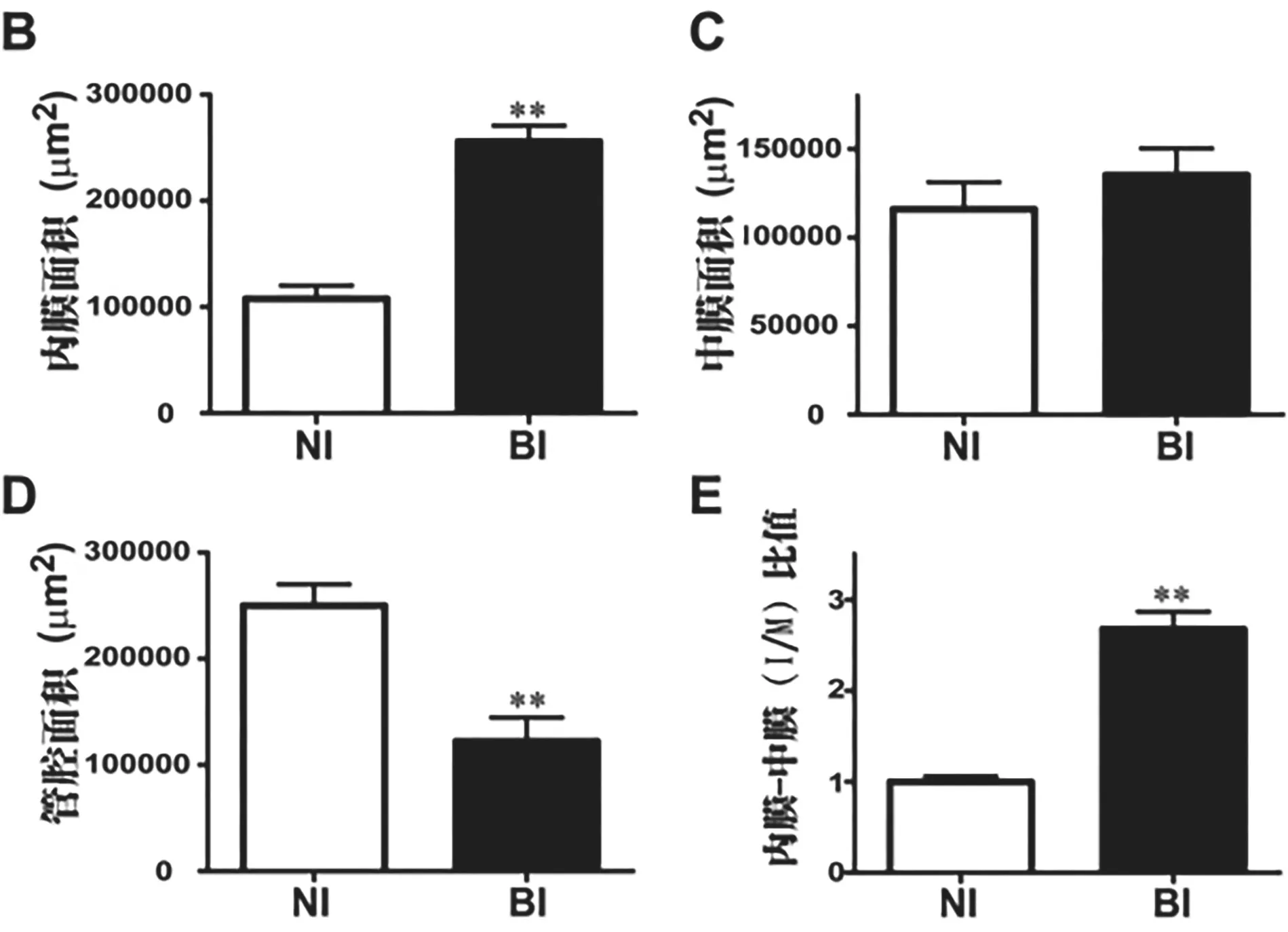
图2 球囊损伤促进新生内膜形成情况。(A)颈总动脉横截面的典型H&E染色显微照片。L:管腔;M:中膜;I:内膜;N:新生内膜。(B)内膜面积的定量分析。(C)中膜面积的定量分析。(D)管腔面积的定量分析。(E)内膜与中膜(I/M)比值的定量。★★P<0.01
2.3 ERK和Akt表达上调 ERK途径和PI3K/Akt途径被证实是新生内膜形成的关键。在本研究中,总ERK、p-ERK、总Akt和p-Akt在BI大鼠中呈上调表达(见图3A),与NI大鼠相比,Akt和ERK的磷酸化比率也呈上调表达(见图3B,P<0.01)。由此推测,Cx37和Cx43的表达可能通过ERK和Akt通路调节。
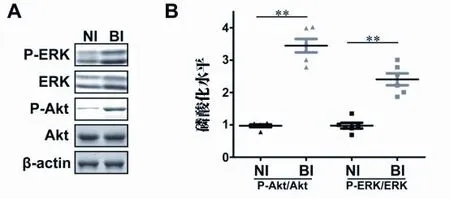
图3 ERK和Akt的磷酸化水平。(A)Akt和ERK磷酸化水平的Western印迹分析,以β-肌动蛋白为对照。(B)Akt和ERK磷酸化水平的量化。通过比较Akt和ERK的磷酸化比率,可以发现球囊损伤组磷酸化水平显著升高。★★P<0.01。
3 讨论
连接蛋白是缝隙连接的组成蛋白,而缝隙连接是一簇连接两个细胞的通道,由连接蛋白体和半通道组成,分别来自于相连接的两个细胞。这种细胞间的通道能够促进化学和电信号的传播,对于血管的功能具有重要作用[4]。在21种已经鉴定的连接蛋白中,血管内皮细胞主要表达Cx37、Cx40和Cx43[5]。
在本研究中,Cx37和Cx43均参与了球囊损伤引起的内膜增生,其机制可能与ERK和Akt通路的调节相关。实验证明,机械损伤诱导的生长因子释放可激活ERK级联,使缝隙连接蛋白能参与组装、分解、运输和降解的调节,以及缝隙连接通道的门控[6]。而Akt在内膜增生过程中的作用则较少被讨论。本研究提示ERK和Akt均参与调解内膜增生,但具体机制尚需进一步研究。
连接蛋白在不同组织中的不同表达模式表明,通过缝隙连接可能有分子运动特异性。值得注意的是,Cx40/Cx43的表达率可以影响异质/异型缝隙连接通道的特性。此外,连接蛋白的磷酸化可以调节连接通透性[7]。Cx37和Cx43的共表达、相互表达或磷酸化调节是否参与球囊损伤诱导的内膜增生尚需进一步研究。
静止的血管平滑肌细胞重新进入细胞周期,进行增殖和迁移是内膜增生过程中的标志性生物学事件,通常认为这是由血管平滑肌细胞与内皮细胞之间的缝隙连接通讯所调控的。因此,有必要深化对连接蛋白调控的研究。
虽然球囊损伤诱导内膜增生的具体机制尚不清楚,但本研究提示ERK和Akt介导的下调Cx37和上调Cx43参与了这一过程。总之,球囊损伤诱导的内膜增生过程中,连接蛋白Cx37下调,Cx43上调,其机制可能与ERK和Akt的介导相关。
