长期氮、磷添加对青藏高原2种高寒草甸植物光合特性的影响
2019-10-25王朋朋王丹王昊
王朋朋 王丹 王昊

摘要:2015、2016年在中国科学院海北高寒草地生态系统国家野外科学观测研究海北站,选取矮蒿草中2种典型的优势物种披碱草(Elymus dahuricus)和麻花艽(Gentiana straminea)为材料,以不添加氮(N)、磷(P)为对照(CK),研究每年分别添加N 10 g/m2、P 5 g/m2及其NP合施对披碱草、麻花艽光合指标的影响。结果表明,添加N,2015、2016年披碱草叶片的净光合速率、蒸腾速率、气孔导度、水分利用效率均小于CK,2015年,麻花艽的净光合速率、蒸腾速率、气孔导度、胞间CO2浓度高于CK;添加NP,2015、2016年,2种高寒草甸植物的净光合作用速率、气孔导度、水分利用效率较CK多有不同程度的增加;单纯添加P,披碱草的表观量子效率较CK升高,单纯添加N,麻花艽的光饱和点较CK升高,而添加NP,披碱草的光补偿点、最大光合速率较CK增加;添加N、P、NP,2种高寒草甸植物叶片的最大羧化速率(Vcmax)、RuBP再生能力(Jmax)较CK多有不同程度的增加,麻花艽的Vcmax值高于披碱草。
关键词:青藏高原;高寒草甸;披碱草;麻花艽;氮;磷;光合特性
中图分类号:S181 文献标志码: A 文章编号:1002-1302(2019)13-0325-05
自工业革命以来,化石燃料的大量使用导致全球CO2等温室气体大量排放,产生温室效应等一系列影响全球生态的问题[1]。全球每年因人类活动向大气中排放的碳约8.7×109 t,而每年大气中碳的增加量不到其50%,生物圈碳库对碳平衡的调节起着重要的作用,生态系统碳循环成为各国学者关注的热点问题[2],而人类活动导致大气氮(N)沉降量成比例增加,明显改变了陆地生态系统的碳、氮循环。我国境内大气干湿沉降总量已成为继欧洲、北美之后的第三大氮沉降区,青藏高原地区大气氮沉降的量较其他地区高[3],而氮沉降输入明显改变陆地生态系统的碳、氮循环速率,进而影响整个生态系统的结构和功能。生态系统几乎都受到N、磷(P)或NP的同时限制[4],生态系统中施加氮、磷通常会促进植物的生长,或出现N饱和后可能受其他养分限制而抑制植物生长的现象[3]。
草地生态系统占全球陆地面积的1/5左右,净初级生产力约占全球陆地生物总净初级生产力的1/3[5]。青藏高原高寒草甸是世界面积最大、海拔最高的草地生态系统,在全球碳、氮循环过程中扮演着重要的角色[6],由于其所处自然环境寒冷潮湿,土壤中的有机氮、磷矿化作用比较低,高寒草甸的植物生長很可能受到氮、磷供应不足的限制[7]。光合作用是绿色植物利用光合色素等将无机物转化为有机物而形成初级生产量的生理过程,养分可对植物叶绿体和光合酶产生影响,进而影响植物的光合作用和干物质累积[8]。
近年来,有关气候变化对高寒草甸影响的研究主要集中在土壤、生产力及植物群落结构上[9-14],基于以大气氮沉降为主要特征气候变化,对高寒草甸植物叶片层次的光合作用、气孔尺度上的生理生态响应与适应策略等方面的研究相对较少。本试验通过探讨不同养分添加条件下高寒草甸植物生理特性的变化规律,研究青藏高原典型高寒植物对不同氮、磷添加条件下的光合生理响应,这将有助于了解高寒草甸植物对气候变化的生理响应机制,进而制定科学的应对方法,确保青藏高原高寒草甸系统的可持续发展。
1 材料与方法
1.1 试验设计
试验于2015—2016年进行,试验地位于中国科学院海北高寒草地生态系统国家野外科学观测研究海北站,37°37′N,101°12′E,选择当地牧民春季或者冬季放牧的牧草地为试验样地,为消除动物粪便对试验的影响,在试验样地四周增加围栏,并捡除样地内可见的所有动物粪便。依据海北站每年实际的大气氮沉降量[3],试验共设计4个养分添加处理,分别为每年添加N 10 g/m2,编号为N;添加P 5 g/m2,编号为P;N、P同时添加,用量分别为10、5 g/m2,编号为NP;不采取任何施肥措施[7],为空白对照,编号为CK。N以尿素的形式添加,P以Ca(H2PO4)2·H2O的形式添加,将预先称好的颗粒状肥料一次性均匀撒在试验样方内。为保证施肥效果,施肥时间选择高寒草甸植物生长季中期(6—8月)的一个雨天或晴天傍晚进行,雨水或次日形成的露水可使肥料迅速溶解进入土壤。每处理重复6次,共计24个样方,每个样方面积为 3 m×3 m。
1.2 光响应参数的测定
每个试验样方内,选取矮蒿草中2种典型的优势物种披碱草(Elymus dahuricus)、麻花艽(Gentiana straminea)健康、阳面叶片为对象,采用美国产LI-6400便携式光合测定仪测量叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间二氧化碳浓度(Ci)、光合有效辐射(PAR)等光合指标,至少重复5次。光合测定仪控制条件为:气室内CO2浓度为 400 μmol/mol,叶室内部光照度为1 500 μmol/(m2·s)、温度为25 ℃,水分吸收装置相对湿度保持在50%~60%。用荧光叶室夹住选好的叶片,保证叶片完全覆盖叶室中间的圆孔,待相应数值稳定后便开始测量相应光合指标。
Pn-Ci曲线:控制CO2浓度以400、300、40、75、150、250、400、800、1 000 μmol/mol梯度变化,保持其他光合测定仪控制条件,测量相应光合指标,并制得相应曲线。Pn-PAR曲线:同理,控制光照度以2 000、1 500、800、300、150、90、40、0 μmol/(m2·s) 梯度变化,保持其他光合测定仪控制条件,测量相应光合指标,并制得相应曲线。
在此基础上,以CO2浓度为400 μmol/mol、光照度为 2 000 μmol/(m2·s)时测得的相应光合指标作为用于试验分析的瞬时气体交换指标,计算水分利用效率(WUE),公式为:
WUE=Pn/Tr。
同时,根据Farquhar等的经验模型FvCB模型[15],估算不同N、P处理下植物叶片的最大羧化速率(vcmax)、RuBP再生能力(Jmax);利用Photosyn Assistant软件得出光补偿点(LCP))、光饱和点(LSP)、最大净光合速率(Pmax)及表观量子效率(EAQ)。
1.3 数据统计分析
采用Excel 2010、SPSS 22.0软件对试验数据进行统计分析和作图。
2 结果与分析
2.1 氮、磷添加对高寒草甸植物瞬时气体交换指标的影响
2.1.1 净光合速率(Pn) 由图1可见,2015年,添加P、NP的披碱草Pn与CK相比有所升高,分别较CK增加41.60%、34.46%,相互间差异不显著(P>0.05);2016年,添加P的Pn较CK显著增加76.38%(P<0.05),添加NP的较CK增加42.96%;2015、2016年,添加N的披碱草Pn较CK均有所降低;2015年,各处理对麻花艽Pn的影响较小,相互间差异不显著,2016年,添加N、NP的麻花艽Pn分别比对照显著降低24.76%、32.90%。
2.1.2 蒸腾速率(Tr) 植物蒸腾是“土壤-植物-大气”水热传输过程中一个极为重要的环节,是植物和环境之间物质交换和能量平衡的重要生理过程,是植物吸收矿物质、光合作用、呼吸作用等生理活动必要的代谢基础。由图1可见,2015年,与CK相比,添加N、P、NP处理对披碱草、麻花艽Tr的影响不显著(P>0.05);2016年,添加P的披碱草Tr较CK显著增加49.90%(P<0.05),添加NP的较CK增加12.04%,与CK相比差异不显著,添加N、NP的麻花艽Tr分别较CK显著减小31.91%、39.24%。
2.1.3 气孔导度(Gs) 气孔是植物与外界环境进行水汽交换的通道,光合作用原料及蒸腾作用的产物进出都要通过气孔,因此,气孔张开闭合的程度会对植物的生理过程产生重要影响[16]。由图1可见,N、P添加对2种植物Gs有明显的影响,麻花艽较披碱草有较高的Gs;2015年,与CK相比,添加N、P、NP处理对披碱草、麻花艽Gs的影响不显著(P>0.05);2016年,添加P的披碱草Gs较CK增加48.81%(P<0.05),添加N、NP的麻花艽分别较CK显著降低34.11%、46.15%。
2.1.4 水分利用效率(WUE) 水分利用效率一般指植物消耗单位质量的水所制造的干物质的量,通常认为是植物在胁迫下产量的一个重要决定因素[17]。由图1可见,2015、2016年,添加N的披碱草水分利用效率(WUE)相对最低,添加NP的披碱草WUE相对较高;2015年,添加NP的披碱草WUE较CK增加6.82%,與CK相比差异不显著(P>0.05);2016年,添加NP的披碱草WUE较CK显著增加28.00%(P<0.05),添加P的麻花艽WUE较CK降低4.09%,与CK相比差异不显著。
2.1.5 胞间二氧化碳浓度(Ci) 胞间二氧化碳浓度可间接反映羧化酶羧化点的浓度,对植物光合作用具有重要意义[18]。胞间二氧化碳浓度与大气二氧化碳浓度比是一个重要的植物生理生态值,不仅与叶片光合羧化酶有关,也与叶片气孔的调节、所处的环境因子有关。由图1可见,2015、2016年,添加N的披碱草Ci相对最大,分别为302.00、291.94 μmol/mol,分别较CK增加2.67%、1.19%,添加NP的Ci相对最小,分别为288.95、261.22 μmol/mol;2015、2016年,添加N、P、NP的麻花艽Ci与CK相比差异不显著(P>0.05)。
2.2 氮、磷添加对光合作用光响应曲线参数的影响
2.2.1 表观量子效率(AQE) 由表1可见,各处理组的AQE相互间差异不显著(P>0.05);2015年添加P的披碱草、2016年添加P的麻花艽、披碱草其表观量子效率(AQE)相对最大,分别较CK增加25.00%和133.33%、10.26%;除2015年的披碱草外,其他处理组均表现为添加N的AQE相对最小。
2.2.2 光饱和点(LSP) 由表1可见,2015、2016年,添加NP的披碱草其光饱和点(LSP)相对最大,分别较CK增加34.64%、31.95%,与CK相比差异不显著(P>0.05);2015、2016年,添加N的麻花艽LSP相对最大,分别较CK增加 5.18%、6.18%,同样与CK相比差异不显著;2015年,添加N的披碱草LSP较CK明显减小37.44%;2016年,添加NP的麻花艽LSP较CK减小32.53%。
2.2.3 光补偿点(LCP) 由表1可见,2015、2016年,添加N的披碱草光补偿点(LCP)相对最大,分别较CK增加 93.28%、142.99%,添加的麻花艽LCP相对最小,分别较CK减小66.64%、10.72%,与CK相比差异不显著(P>0.05)。
2.2.4 最大光合速率(Pmax) 由表1可见,2015、2016年,添加NP的披碱草最大光合速率(Pmax)相对最高,分别较CK增加38.82%、114.42%,添加N的相对最低,分别较CK减小21.38%、19.30%,与CK相比差异不显著(P>0.05);2015、2016年,CK处理的麻花艽Pmax相对最高,不同添加N、P、NP的麻花艽Pmax均有不同程度的降低,其中,2015年添加N、P、NP的麻花艽Pmax较CK有显著降低(P<0.05),分别较CK减小 33.72%、32.43%、44.21%。
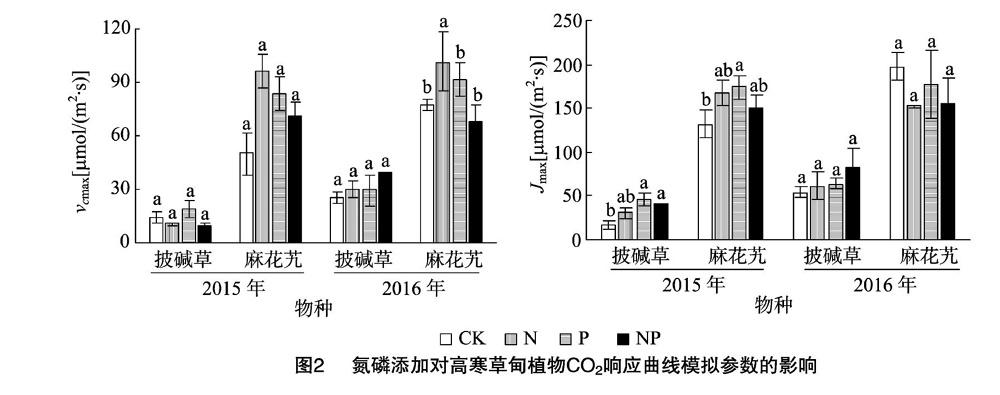
2.3 氮、磷添加对高寒草甸植物FvCB模型参数的影响
FvCB模型是解释和模拟叶片气体交换中使用最为广泛的一个模型[15]。由图2可知,披碱草、麻花艽叶片最大羧化速率(vcmax)对添加N、P的响应并不相同,有较为明显的差异,2015、2016年麻花艽的vcmax值均高于披碱草;2016年,添加N、P、NP的披碱草vcmax值较CK均有所增加,分别比CK增加19.73%、16.50%、56.00%;2015年,增加N、P、NP的麻花艽vcmax值分别较CK增加93.34%、67.80%、42.24%,2016年,添加N、P处理的麻花艽vcmax值分别较对照增加31.47%、18.19%,且添加N的显著高于CK(P<0.05),但添加NP的麻花艽vcmax值明显低于CK。
对RuBP再生能力(Jmax)而言,2015、2016年,添加N、P、NP的披碱草Jmax较CK均有所增加,分别比CK增加 71.98%、154.20%、129.11%和13.83%、18.14%、52.42%,2015年添加P、NP处理的显著高于CK;2015年,添加N、P、NP的麻花艽Jmax较CK均有所增加,分别比CK增加 26.11%、31.58%、12.94%,其中,添加P的处理显著高于CK;2016年,添加N、P、NP的麻花艽Jmax均呈现不同程度的降低,但相互间差异不显著(P>0.05)。
3 结论与讨论
有研究发现,氮(N)添加对植物的净光合速率(Pn)可起到一定的促进作用[19-21],而高N条件下,作物的光合作用可能被抑制[22]。本研究发现,在光合、叶片性状存在差异的披碱草、麻花艽这2种高寒植物,2015、2016年添加N的披碱草和2016年添加N的麻花艽其净光合速率均有所降低,而2015、2016年添加NP的披碱草Pn有所增加,可能是由于高寒草甸植物吸收的N元素形成NH4+,扰乱了植物细胞内N、P等元素的代谢,从而使Pn降低,也可能是由于叶片中增加的N没有被分配给RuBP羧化酶(Rubisco)或叶绿素等,并被用于植物光合上。N对植物生长的影响主要表现在光合作用和促进叶片生长2个方面[23],而本研究中N的添加并未明显促进植物的光合作用,可能是长期高氮处理消除了对光合性能的氮限制,进而受到其他营养元素的限制[24]。
磷(P)是多种植物功能所必需的植物养分,是植物体内核酸、腺嘌呤核苷三磷酸(ATP)、磷脂等的主要成分,而這些物质在植物光合中起到重要的作用[25]。P限制植物的光合作用,是由于P缺失会降低植物RuBP酶的再生[26],导致光能利用效率和气孔导度的下降[27-28]。P不仅自身会影响植物的生理特性,还会影响Rubisco酶的活性[29]。有研究发现,P的添加可以显著增加植物的地上生物量[7],会使植物叶片和最大光合作用速率之间的线性关系斜率变大[30]。本研究结果表明,添加P可明显提高2种高寒植物的净光合速率(Pn),添加NP对这2种植物的光合能力没有明显提高作用。这说明在青藏高原高寒生态系统中,2种高寒草甸植物在长期P添加及大气氮沉降影响下,出现氮饱和现象,光合作用不受氮有效性的限制,而P的加入所引起的植物光合作用比N的正反应更强。
本试验结果表明,2015、2016年,添加N的麻花艽光饱和点较CK有所升高,2015年的光补偿点稍有降低,这有利于延长光合时效,提高麻花艽光合产物的积累量;随着P的添加,添加P、NP的2种高寒草甸植物最大光合速率、胞间CO2浓度(Ci)、气孔导度、蒸腾作用速率多有不同程度的升高,说明P能有效促进2种高寒草甸植物的光合作用和蒸腾作用,这与吉艳芝等的研究结果[31]一致。营养物质的添加不仅影响植物的碳积累,而且影响其叶片气孔功能。气孔导度(Gs)是反映气孔行为的重要指标之一,也是重要的光合参数[32]。本研究发现,添加P可明显提高披碱草、麻花艽的Gs;添加N时,2种高寒草甸植物净光合速率、Gs多有不同程度的下降,而Ci多有上升,说明Gs降低可能是叶肉细胞羧化能力降低引起的结果。
Farquhar等发现,环境因子对植物光合作用的影响,首先表现为植物叶片最大羧化速率(vcmax)、RuBP再生能力(Jmax)的改变[15]。正常条件下,植物光合作用Rubisco酶活性越高或含量越大,其vcmax越大,同时植物体内N含量与vcmax相关性很大,植物体内N浓度的增加是引起植物vcmax增大的最主要因素,从而促进作物光合作用增强。本研究发现,添加N、P,高寒草甸植物vcmax、Jmax值多有不同程度的增加,而vcmax和Jmax的增加是植物提高自身光合能力的表现。
总之,本研究以披碱草和麻花艽这2种高寒草甸植物为研究对象,初步掌握了其在不同养分搭配条件下的光合特性变化规律,这可为今后青藏高原高寒草甸的规模化管理提供参考。须指出的是,本研究仅针对不同养分添加条件下高寒草甸植物光合特性的变化进行分析,而没有涉及植株对养分的吸收、利用、转化等生理生化过程,今后研究中应进行更深入探讨,为高寒草甸草地生态系统规模化、优质化管理提供更多的科学依据。
参考文献:
[1]Lin H L,Wang J,Xu Z,et al. Research progress and trend of the carbon cycle in grassland agroecosystem[J]. Pratacultural Science,2005,22(4):59-62.
[2]刘东生. 全球变化和可持续发展科学[J]. 地学前缘,2002,9(1):1-9.
[3]Lu C Q,Tian H Q. Spatial and temporal patterns of nitrogen deposition in China:synthesis of observational data[J]. Journal of Geophysical Research-Atmospheres,2007,112(D22):S05.
[4]Elser J J,Bracken M E,Cleland E E,et al. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater,marine and terrestrial ecosystems[J]. Ecology Letters,2007,10(12):1135-1142.
[5]Saugier B,Roy J,Mooney H A. Terrestrial global productivity:past,present and future[M]. San Diego:Academic Press,2001:89-91.
[6]Scurlock,O J M,Johnson,et al. Estimating net primary productivity from grassland biomass dynamics measurements[J]. Global Change Biology,2002,8:736-753.
[7]杨晓霞,任 飞,周华坤,等. 青藏高原高寒草甸植物群落生物量对氮、磷添加的响应[J]. 植物生态学报,2014,38(2):159-166.
[8]毛晋花,邢亚娟,马宏宇,等. 氮沉降对植物生长的影响研究进展[J]. 中国农学通报,2017,33(29):42-48.
[9]武丹丹,井 新,林 笠,等. 青藏高原高寒草甸土壤无机氮对增温和降水改变的响应[J]. 北京大学学报(自然科学版),2016,52(5):959-966.
[10]潘新丽. 模拟增温对川西亚高山人工云杉林土壤碳含量和土壤呼吸的影响[J]. 生态研究,2008,19(8):1637-1643.
[11]徐满厚,刘 敏,翟大彤,等. 青藏高原高寒草甸生物量动态变化及与环境因子的关系——基于模拟增温实验[J]. 生态学报,2016,36(18):5759-5767.
[12]宗 宁,段 呈,耿守保,等. 增温施氮对高寒草甸生产力及生物量分配的影响[J]. 应用生态学报,2018(1):59-67.
[13]李 娜,王根绪,杨 燕,等. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响[J]. 生态学报,2011,31(4):895-905.
[14]赵建中,刘 伟,周玉碧. 模拟增温对矮嵩草草甸莎草科功能群植物的影响[J]. 青海草业,2012,21(2):2-7,18.
[15]Farquhar G D,von Caemmerer S,Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species[J]. Planta,1980,149(1):78-90.
[16]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiol,1982,33:317-346.
[17]Blum A. Effective use of water(EUW)and not water-use efficiency(WUE)is the target of crop yield improvement under drought stress[J]. Field Crops Research,2009,112(2):119-123.
[18]Farquhar G D. OLeary M H,Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves[J]. Functional Plant Biology,1982,9(2):281-292.
[19]吴 楚,王政权,范志强. 氮素形态处理下水曲柳幼苗养分吸收利用与生长及养分分配与生物量分配的关系[J]. 生态学报,2005,25(6):1282-1290.
[20]Lebauer D S,Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed[J]. Ecology,2008,89(2):371-379.
[21]李德軍,莫江明,方运霆,等. 模拟氮沉降对三种南亚热带树苗生长和光合作用的影响[J]. 生态学报,2004,24(5):876-882.
[22]翟占伟,龚吉蕊,罗亲普,等. 氮添加对内蒙古温带草原羊草光合特性的影响[J]. 植物生态学报,2017,41(2):196-208.
[23]Gastal F,Lemaire G. N uptake and distribution in crops:an agronomical and ecophysiological perspective[J]. Journal of Experimental Botany,2002,53(370):789-799.
[24]Tang Y L,Wen X G,Lu Q T,et al. Heat stress induces an aggregation of the light-harvesting complex of photosystem Ⅱ in spinach plants[J]. Plant Physiology,2007,143(2):629-638.
[25]Benon H J,Bielski,Chan P C. Enzyme-catalyzed free radical reactions with nicotinamide-adenine nucleotides:Ⅰ.Lactate dehydrogenase-catalyzed chain oxidation of bound NADH by superoxide radicals[J]. Archives of Biochemistry and Biophysics,1973,159(2):873-879.
[26]Jacob J,Lawlor D W. Dependence of photosynthesis of sunflower and maize leaves on phosphate supply,ribulose-1,5-bisphosphate carboxylase/oxygenase activity,and ribulose-1,5-bisphosphate pool size[J]. Plant Physiology,1992,98(3):801-807.
[27]Conroy J P,Smillie R M,Kuppers M. Chlorophyll a fluorescence and photosynthetic and growth responses of pinus radiata to phosphorus deficiency,drought stress,and high CO2[J]. Plant Physiology,1986,81(2):423-429.
[28]Jacob J,Lawlor D W. Extreme phosphate deficiency decreases the in vivo CO2/O2 specificity factor of ribulose 1,5-Bisphosphate carboxylase-oxygenase in intact leaves of sunflower[J]. Journal of Experimental Botany,1993,44(268):1635-1641.
[29]Warren C R,Adams P R. Uptake of nitrate,ammonium and glycine by plants of Tasmanian wet eucalypt forests[J]. Tree Physiology,2007,27(3):413-419.
[30]Reich P B,Oleksyn J,Wright I J. Leaf phosphorus influences the photosynthesis-nitrogen relation:a cross-biome analysis of 314 species[J]. Oecologia,2009,160(2):207-212.
[31]吉艷芝,陈立新,薛宝民,等. 施肥对落叶松人工林植物养分及生理特性的影响[J]. 生态环境,2004,13(2):217-219.
[32]徐振锋,胡庭兴,张 力,等. 青藏高原东缘林线交错带糙皮桦幼苗光合特性对模拟增温的短期响应[J]. 植物生态学报,2010,34(3):263-270.
