杂交鹅掌楸的生长调查与家系选择
2019-10-25肖保荣吴伟煌黄世清
肖保荣 吴伟煌 黄世清

摘要:为了评价不同交配组合F1代杂交鹅掌楸(Hybrid liriodendron)家系在湖北地区的生长适应情况,选育适合我国中部地区推广应用的优良杂交鹅掌楸品系,以6株中国鹅掌楸为母本,4株北美鹅掌楸为父本,进行人工控制授粉,共获得21个杂交子代家系。于2014年将其中19个子代家系和1个中国鹅掌楸混系(对照)1年生幼苗定植于湖北省黄冈市林场,分别在2年和3年后进行生长测定。2016年测量各子代家系树高(height)和米径(diameter)2个指标,2017年测量各子代家系树高(height)和胸径(DBH)2个指标,进行表型性状和遗传参数分析。结果表明,家系16、5、10、15在树高、米径和胸径性状均值上都具有显著优势;家系20、17、16、5、4的树高、米径和胸径性状的变异系数明显低于其他家系;家系9、12、20、4的树高、米径和胸径性状遗传力明显高于其他家系;家系16、4、3、19的育种值显著高于其他家系。可见,在筛选优势生长的家系方面,树高、米径和胸径的平均数值都比较高的家系,有更好的用材价值和经济价值;家系的数量性状变异系数越小,生长越稳定,时间上的变异较小;遗传力和育种值越大,家系的遗传力越稳定,更加适合子代育种的选择;杂交的亲本树龄适宜选择母本为13年生的中国鹅掌楸和父本为22年生的北美鹅掌楸进行杂交,F1代的遗传力较好并且育种值较高,适宜推广种植。
关键词:杂交鹅掌楸;遗传力;遗传变异;育种值;家系选择
中图分类号:S792.210.4 文献标志码: A 文章编号:1002-1302(2019)13-0183-05
杂交育种是指通过不同的种间树种进行交配获得的子代的过程。英国学者Henry在1912年开展了杨树(Populus)、白蜡树(Fraxinus chinensis Roxb.)和栎类(Quercus)等种间杂交,第一次在林木改良中提出雜交的概念,并且成为育成杨树人工杂交品种的第一人[1]。我国的林木杂交工作开始于1948年,由已故著名林木遗传育种学家叶培忠教授进行杨树杂交的初步试验,取得了很好的成就。而大规模杂交育种是从1955年开始,以杨树人工杂交为起点,开展了杉(Taxodiaceae)、松(Pinus)、板栗(Castanea mollissima)等树种的杂交育种[2]。杂交鹅掌楸(Hybrid liriodendron),又名杂交马褂木,为鹅掌楸属种间杂交种,具有很高的可配性和明显的生长优势[3],上个世纪70年代由著名的林木育种学家叶培忠教授首次培育成功,杂交鹅掌楸材质优良,其叶形似马褂奇特、整体树形蜿蜒优美、花朵大且颜色鲜艳,而且适应性强、生长快速,具有广阔的园林绿化和用材林营造等应用前景[4]。
杂交鹅掌楸是中国和北美鹅掌楸种间杂交树种,具备北美鹅掌楸的快速生长和中国鹅掌楸对环境灵敏度高的特性。王章荣等的研究结果显示,杂交鹅掌楸20年生时树高、胸径、材积的杂种优势分别达到20.79%、69.59%、232.13%[5]。杂交育种是植物遗传改良的一种有效手段和主要途径,是目前最有成效的育种方法之一,在杂交鹅掌楸家系生长性状的遗传变异方面,李周崎等研究表明杂交鹅掌楸的苗高性状,家系平均变异系数为29.16%、家系内平均变幅为2.31 m,在地径性状上,家系平均变异系数为31.87%、家系内平均变幅为2.63 cm[6]。张武兆等对15年生鹅掌楸杂交子代生长测定结果表明,鹅掌楸杂种优势非常明显,胸径年生长量达2 cm,树高年生长量1 m以上[7]。李火根等研究认为,北美鹅掌楸胸径、树高早期增速较快,进入中壮龄阶段竞争加剧,个体间分化增大[8]。李斌等研究认为,中国鹅掌楸生长性状在种源间存在显著的遗传差异。地点间差异极显著,种源对环境反映灵敏,种源与地点间存在明显的交互作用[9]。刘洪谬等研究表明,与中国鹅掌楸相比,杂交鹅掌楸12年时树高、胸径和材积的杂种优势分别达到35%~53%、30%~83%、129%~411%[10]。张晓飞等研究发现,无论是苗期还是幼龄期,鹅掌揪不同交配组合子代的生长量均存在显著的基因型环境交互作用,不同组合的遗传稳定性差异显著[11]。杂交鹅掌楸子代家系的遗传变异丰富,对环境响应差异明显,基于杂交鹅掌楸家系间的变异进行优良家系选育,必要而且可行。
杂交鹅掌楸是我国南方低山丘陵地区理想的造林树种,目前已在福建、江苏、浙江、江西和湖南等地广泛引种栽植,长势良好[12-16]。但目前在我国中部地区有关杂交鹅掌楸生长适应性研究和报道仍较少,对杂交鹅掌楸在该部地域推广应用的科学性评价仍相对较薄弱。因此,本研究以18个杂交鹅掌楸子代家系和1个中国马褂木混系为研究对象,综合评价杂交鹅掌楸在黄冈地区的适应性,以期初步选育出若干长势优异、稳定性好的杂交鹅掌楸家系,为杂交鹅掌楸在我国中部地区的进一步推广和应用,争取营林效益的最大化奠定理论基础。
1 材料与方法
1.1 研究材料
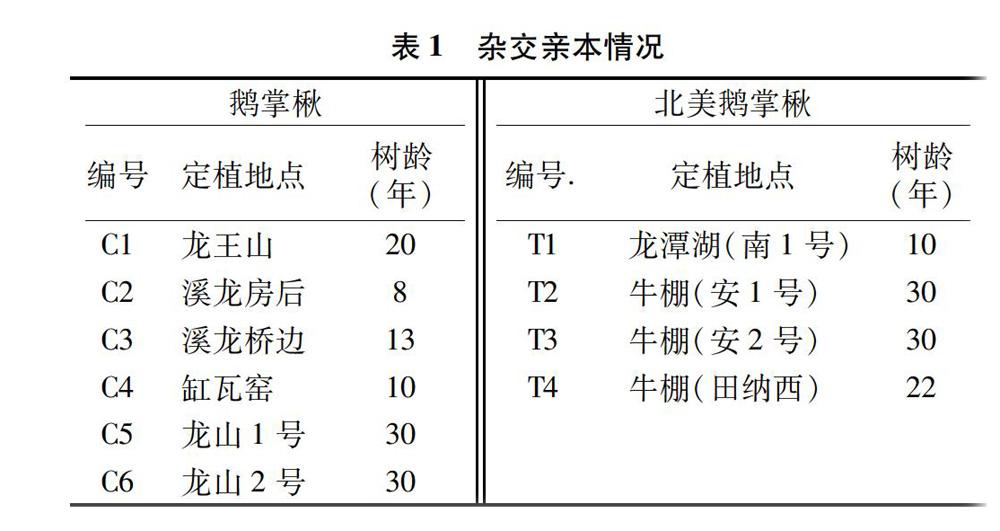
2012年以6株马褂木作为母本、4株北美鹅掌楸作为父本开展杂交育种(表1),共21个组合。10月初采种。次年春季育苗。2014年春季,将其中19个杂交鹅掌楸子代家系及1个中国鹅掌楸混系(对照)1年生苗定植于湖北黄冈,由于1号和21号苗数量较少,未种植,株行距为3 m×4 m,每个家系每个重复12株,共设4个重复。试验地位于湖北省东部,大别山南麓,长江中游北岸,114°55′E,30°27′N,海拔 25 m。地区气候属亚热带季风气候,日照充足,雨量充沛,四季分明。年均日照时数为2 082 h,年均气温为17 ℃左右,年均降雨日为100.1 d,降水量为1 213.4 mm,年均无霜期为261 d,初霜日始于11月24日至12月1日,霜期止于3月 8—16日。土壤为江河冲击平原,属灰潮土类型,土质为沙壤土,pH值7.5,立地条件较好,原为耕地,土层深厚,较肥沃[17]。立地条件适宜杂交鹅掌楸的生长。
1.2 杂交母树及杂交家系的鉴定
以笔者所在实验室自主开发的引物对杂交母树及杂交后代进行鉴定,引物序列及鉴定方法参考龙晓飞等的方法[18]。
1.3 数据统计及分析方法
分别在2016年6月和2017年6月对试验地各子代家系每株树高(height)、米径(diameter)胸径(DBH)、进行生长测定,使用SPSS 21.0软件进行方差分析和均方分析,由于家系8没有存活,所以没有测量,结合ASReml统计软件,利用最佳线性无偏估计(BLUE,best linear unbiased estimation)方法得到固定效应的最佳无偏估计值和利用最佳线性无偏预测(BLUP,best linear unbiased prediction)方法计算得到估计遗传力值,利用R语言进行家系的遗传力估计和Excel表格作图。
2 结果与分析
2.1 不同家系苗期生长差异比较分析
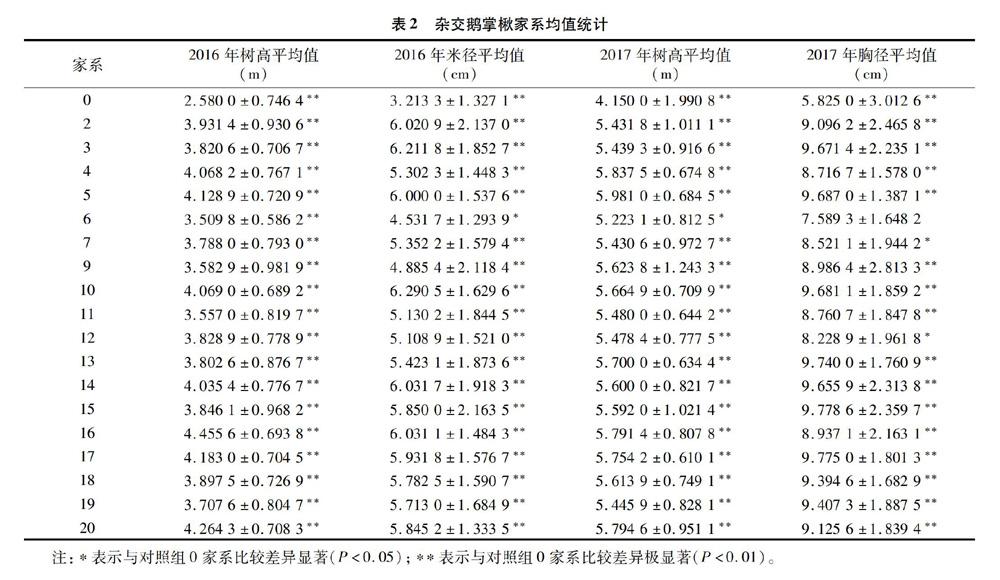
连续2年对19个定植3年的杂交鹅掌楸家系的树高和胸径进行均值和标准差统计分析,结果见表2和图1。整体上看,2016年所有家系与家系0的树高性状对比差异性都是极显著的,家系6的米径性状在2016年差异性和2017年树高性状的差异性均表现为显著,而在2017年胸径性状的差异性表现为不显著。家系7和12的2017年胸径性状的差异性表现为显著,其余家系的2个性状与家系0比较的差异性均表现为极显著。
从图1整体均值的趋势上看,19个家系连续2年都是米径和胸径的变化明显,2016年和2017年树高均值最大的是
从遗传力的分析图(图3)可以得知,遗传力相对于树高和胸径整体波动较大。同一个家系在连续2年都出现一定范围的波动,相对波动较小且遗传力都维持在一个较高值的是家系2、9、20。从遗传的稳定性考虑,这3个家系适宜作为优势树种选择。
2.4 杂交亲本组合确定
综合遗传力和育种值分析,优势的家系分别是家系2、3、4、9、12、16、20,对应的亲本杂交组合(表5)分别是♀C2×♂T1、♀C2×♂T2、♀C2×♂T3、♀C3×♂T4、♀C4×♂T3、♀C4×♂T3、♀C6×♂T3这7个。
3 结论与讨论
在均值比较方面,通过连续2年对19个不同家系树高和胸径的均值比较分析,仅按照均值最大的标准分别筛选出优势的家系有家系16、5、10、15,這4个家系可以作为优势参考树种在黄冈地区推广。杂交鹅掌楸具有良好的景观价值,树形美观且冠幅大叶形奇特,可以优先考虑在湖北省的风景区进行种植推广。
在性状变异分析方面,通过连续2年对19个不同家系2个数量性状的变异系数比较分析得知,变异系数越小,性状越稳定,进而筛选出家系20、17、16、5、4,在针对树种家系性状的选择时可以参考性状变异系数分析结果,良好稳定的性状可以作为种植参考依据进一步推广,这将有利于杂交鹅掌楸更好地适应不同的生长环境。
在遗传力分析方面,通过连续2年对19个不同家系F1代的遗传力估计分析得出,家系9的遗传力最高,其次是家系12、20、4,这4个家系可以作为黄冈地区的遗传优势家系进行筛选,遗传稳定性好,也可以作为湖北省地区的杂交鹅掌楸的遗传优势家系进行推广。针对育种值进行分析,筛选出家系16、4、3、19这4个育种值高的家系。
综合性状均值分析,性状稳定性分析、遗传力估计和育种值预测分析,筛选出了家系4、9、10、12、15、16、17、20,并根据这8个家系的亲本组合,分别筛选出♀C2×♂T3、♀C3×♂T4、 ♀C4×♂T1、 ♀C4×♂T3、♀C5×♂T2、♀C5×♂T3、♀C5×♂T4、♀C6×♂T3,这8个亲本组合杂交的子代生长性状优越,子代的性状稳定,遗传力高,育种值大。在树龄选择方面,中国马褂木可以适宜选择8、10、13、30年这4个阶段,北美鹅掌楸适宜选择10、22、30年这3个阶段,杂交子代的生长性状稳定性良好,遗传力和育种值高。
综上所述,在湖北地区进行杂交马褂木的家系推广,宜选择家系4、9、10、12、15、16、17、20这8个优良家系,抗寒能力强,性状的稳定性好,生长适应性良好,遗传力和育种值高,适合推广种植。
4 展望
通过对黄冈地区的连续2年的19个不同家系的杂交鹅掌楸的树高和胸径进行测量并且分析生长状况,初步确定了黄冈地区的优势家系及其遗传稳定性和育种能力,鉴定了杂交亲本的组合方式和树龄,为杂交鹅掌楸在湖北地区的推广提供一定的理论参考依据。
参考文献:
[1]李善文,张志毅,何承忠,等. 中国杨树杂交育种研究进展[J]. 世界林业研究,2004,17(2):37-41.
[2]袁晓峰. 鹅掌楸杂交组合选择及无性系苗期测定[D]. 南京:南京林业大学,2012.
[3]王章荣. 鹅掌楸属(Liriodendron)杂交育种回顾与展望[J]. 南京林业大学学报(自然科学版),2003,27(3):76-78.
[4]向其柏,王章荣. 杂交马褂木的新名称——亚美马褂木[J]. 南京林业大学学报(自然科学版),2012,36(2):1-2.
[5]王章荣. 中国马褂木遗传资源的保存与杂交育种前景[J]. 林业科技通讯,1997(9):6-8.
[6]李周岐,王章荣. 鹅掌楸属种间杂种苗期生长性状的遗传变异与优良遗传型选择[J]. 西北林学院学报,2001,16(2):5-9.
[7]张武兆,马山林,邢纪达,等. 马褂木不同家系生长动态及杂种优势对比试验[J]. 林业科技开发,1997,11(2):32-33.
[8]郝自远,李火根,康 昊,等. 北美鹅掌楸人工林生长规律及早期选择可行性探究[J]. 林业科学研究,2017,30(5):878-885.
[9]李 斌,顾万春,夏良放,等. 鹅掌楸种源遗传变异和选择评价[J]. 林业科学研究,2001,14(3):237-243.
[10]刘洪谔,沈湘林,曾玉亮. 中国鹅掌楸、美国鹅掌楸及其杂种在形态和生长性状上的遗传变异[J]. 浙江林业科技,1991(5):18-22.
[11]张晓飞. 鹅掌楸不同交配组合子代幼龄期生长性状的遗传分析[D]. 南京:南京林业大学,2008.
[12]蔡伟建,杨丽芳,高捍东,等. 杂交马褂木人工林生长规律及趋势研究[J]. 安徽农业科学,2011(15):9457-9459.
[13]严珍花. 杂交马褂木营种造林技术应用及效益初探[D]. 福州:福建农林大学,2015.
[14]颜立红,左海松,殷元良,等. 杂交鹅掌楸丘陵区栽培试验研究[J]. 湖南林业科技,2002(4):19-34.
[15]刘敬优. 坡位和造林密度对杂种马褂木生长的影响[J]. 安徽农学通报,2013(10):87-89.
[16]方禄明. 体细胞胚胎发生杂种马褂木造林密度试验[J]. 林业科技开发,2009(3):99-101.
[17]汪高明. 湖北省近47年气温和降水气候特征分析[D]. 兰州:兰州大学,2009.
[18]陈金慧,龙晓飞,施季森,等. 一种杂交鹅掌楸遗传关系鉴定的方法[P]. 2018-05-29.
