典型高寒植物有性繁殖对长期增温的响应
2019-05-31康濒月徐恒康姜风岩邵新庆
康濒月,徐恒康,姜风岩,邵新庆
(中国农业大学草业科学与技术学院,北京 100193)
全球气候对陆地生态系统的影响及其反馈是全球变化与陆地生态系统(Global Change and Terrestrial Ecosystem,GCTE)研究的热点,据大气环流模型(General Circulation Model,GCM)预测,21世纪全球温度将升高1.5℃~4.5℃[1]。这种全球气候变暖趋势不但会影响植物生理生态特征,也会对植物种群、群落、生态系统乃至整个生物圈产生巨大影响。尤其在高纬度和高海拔的青藏高原,生态环境脆弱[2-3],植物对温度升高的响应非常敏感[4-5]。
有性繁殖是植物繁衍后代的主要方式之一,有利于植物对当地环境的适应,维持遗传多样性[5]。研究表明,有性生殖阶段对环境的变化最敏感[6]。随着全球气候变化,青藏高原气候呈现出暖化趋势,温度升高可能对当地植物生殖生长过程的授粉受精、生殖产量产生明显影响[7]。在研究增温对植物有性生殖的影响中,一些研究者得出了相似的结论:Benlloch-Gonzalez等[8]发现温度升高4℃会降低果实产量并产生较小的果实,也有研究[9-10]表明增温显著降低了短花针茅结实率、种子百粒重和发芽率。但不同研究者通过模拟增温对不同草地生态系统的影响的研究得到的结论不尽相同。曹素珍认为[11]增温降低植物的繁殖投入,促进营养生长,Klady等[12]认为变暖促进了大多数物种的繁殖生长,并且北极地区长期变暖可能会促进苔原植物的有性繁殖;也有研究认为增温对禾草科和杂类草的生长没有显著影响[13]。研究植物繁殖特征对增温的响应,是研究气候变化背景下高寒草甸群落结构演替和功能变化的基础。目前鲜有对植物有性繁殖对增温响应及敏感性的研究,有关这一方面的大部分研究结论大多基于短期和室内控制试验,难以获取高寒植物响应气候变化的综合信息,更难以得出响应敏感性与趋势的结论。因此,本研究在青藏高原海北试验站使用开顶式生长室(Open-Top Chambers,OTCs)进行长期和室外控制模拟增温试验[14-16],以高寒典型植物莎草科矮生嵩草(Kobresiahumilis)、青藏苔草(Carexmoorcroftii)和禾本科垂穗披碱草(Elymusnutans)为研究对象,研究增温处理下这3种植物的结实率、种子产量、种子活力等繁殖特征对增温的响应,分析3种植物的生殖策略,为预测高寒草甸草原植物群落变化趋势以及科学评估气候变化对高寒草甸的影响奠定了数据和理论基础。
1 材料与方法
1.1 研究区概况
本试验在海北高原现代生态畜牧业科技试验示范园进行,该园区位于青海省海北藏族自治州海晏县西海镇,地处青藏高原东北部青海湖东岸(36°55' N,100°57' E,海拔3029 m)。该地区属于高原大陆性季风气候,仅有冷暖两季之分。年均温1.5℃左右,年日照时数2980 h左右,相对湿度为45%~60%,年降雨量约400 mm,降雨主要集中在7-9月。土壤类型为高山草甸土,土层较薄,缺磷,少氮。0~20 cm土壤有机质含量为27.2 g·kg-1,全氮含量2.5 g·kg-1,有效磷含量为0.05 g·kg-1。该区域植被类型为高寒针茅化草甸草,主要优势种为紫花针茅(Stipapurpurea)、草地早熟禾(PoapratensisL.)、冷地早熟禾(Poacrymophil)、垂穗披碱草、线叶嵩草(Kobresiacapillifolia)、矮嵩草、扁蓿豆(Medicagoruthenica)和二裂委陵菜(Potentillabifurca)等。
1.2 试验设计
本试验研究对象为自然状态下的高寒草原化草甸生态系统,自2012年起采用开顶箱(OTCs)式增温装置(图1)对试验样地进行长期增温。根据国际冻原计划(International Tundra Experiment,ITEX)标准建立模拟增温的野外试验平台,试验样地地势平坦,植物分布均匀,用围栏封闭。试验处理分增温(Warming,W)和对照(Control,CK),各8组重复,增温采用OTCs增温装置进行增温2℃,并设计自动控温系统和抽气式风扇,当OTCs内外温差值超过2℃时,风扇自动启动,保证增温不超过设定值,维持温度稳定。OTCs永久性安装于样地里,规格为:底部直径2.2 m,顶部直径1.5 m,高度0.7 m。在7-8月对试验对象:矮生嵩草、青藏苔草和垂穗披碱草进行调查,并在各自结实期进行取样。

图1 开顶式生长室示意图Fig.1 The sketch of the open top chamber
1.3 试验方法
1.3.1土壤温度测定方法 使用AR5数据采集器采集每个样地地上20 cm处的空气温度、湿度和地下5 cm处的土壤温度、湿度(15 min一次)。
1.3.2调查植物种群数量特征测定方法 在草地植物旺盛生长期(7-8月),采用方精云等[19]介绍的样方法对垂穗披碱草、青藏苔草和矮蒿草的种群数量特征进行调查,主要调查指标为相对密度和相对盖度。3种植物种群特征情况见表1。

表1 增温下3种植物种群数量特征Table.1 Population characteristics of three plant species under warming
注:CK:空白对照;W:增温
Note:CK:Control;W:Warming
1.3.3有性繁殖指标测定方法 采用周兴民等[17]介绍的方法,测定草甸植被主要种群繁殖对策参数。具体做法为:在每个样地上布设1 m×1 m样方,2018年8月在3种代表草本植物:矮生嵩草、青藏苔草、垂穗披碱草的各自果期分别对其繁殖枝和营养枝进行计数并采用全收获法[18]收获,将根和根状茎上的泥土洗净、晾干,并将植物构件分为繁殖器官(花、花轴或花茎)和非繁殖器官(茎、叶、根等)和果实三部分,同时记录繁殖枝上的小穗及种子数目。于80 ℃恒温下将植物烘干至恒重(约需要48 h)[19]。用电子天平精确称重各部分质量(精确在0.0001 g),计算有性繁殖效力。有性繁殖效力用穗(包括花和种子)占总生物量(以干重计)的比例来计算。
1.3.4种子生产能力测定方法 在采样过程中基于繁殖枝是否萌发将所有样本分为繁殖性和非繁殖性[20]。对于每株繁殖性植物,对其繁殖枝数进行统计,然后使用枝数来估计每株植物(i)每年(y)产生的种子数量(Seed,S),关系如下:
Si,y=Ti,y×f×s
注:T:繁殖枝数;f:每根繁殖枝上果实数;s:每个果实产生的种子数量
1.3.5种子发芽率测定方法 参考邓自发等[21]介绍的方法采集种子,在各自结实期结束时,在完全随机设计的每个样地内随机采集3种调查植物繁殖枝25枝,逐穗收集种子,对有成熟种子的穗和每穗种子数进行计数。同时为避免OTCs气室的边缘效应,在其中心取样。对样品种子依据完全随机设计试验的样地顺序进行编号并按照标准发芽试验步骤进行室内发芽,采用纸上法在恒温条件下发芽,从每个样地内3种调查植物种子中分别取50粒、4组重复,置于培养皿中润湿的滤纸上(2层),加皿盖在25℃光照处理12 h,暗处理12 h)恒温培养箱中萌发。由于苔草种子的休眠特
性,提前将青藏苔草种子进行低湿层积14 d[22]。萌发期间,保持适宜水分,统计发芽势、发芽率,以胚根突破种皮1 mm或以上计为发芽[23]。14 d结束实验。统计试验末期发芽种子占供试种子的百分数。
发芽率(%)=种子总发芽数/供试种子数×100%
发芽势(%)=规定时间内种子发芽数/供试种子数×100%
1.4 数据分析
采用Excel 2016进行数据整理、计算和作图,数值均以平均值±标准误表示。利用IBM SPSS Statistics 23软件进行单因素方差分析(One-way ANOVA),用LSD法在显著性水平0.05和0.01下进行差异性检验,不同大写字母表示在增温处理下差异极显著(P<0.01),不同小写字母表示增温处理下差异显著(P<0.05)。用R语言3.4.4对3种植物繁殖枝数量、小穗长度、繁殖效力、种子产量、发芽势和发芽率的数据进行heatmap分析并绘制图像。
2 结果分析
2.1 繁殖枝数量、小穗长度和繁殖效力的变化
由图2可知,增温处理下,3种植物繁殖枝数量都有减少的趋势,小穗长度有增加的趋势。其中,矮嵩草繁殖枝数量在增温下极显著降低(P<0.01),平均每株矮嵩草减少1~2枝繁殖枝,其小穗长度的变化趋势与繁殖枝数量变化趋势基本相反(图2),有增长趋势但是与对照之间差异不显著。从图2可见,增温对青藏苔草、垂穗披碱草有效穗长度有显著影响(P<0.05),其中垂穗披碱草有效穗长度在增温处理下平均每株增加2~3 cm,青藏苔草有效穗长度在增温处理下平均每株增加0.5~1 cm。说明在增温影响下,3种调查植物繁殖枝数量与其有效穗长度变化方向相反,且变化幅度不一致。
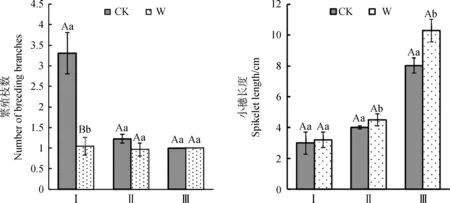
图2 增温对高寒植物繁殖茎数量、小穗长度的影响Fig.2 Effects of warming on the numbers of stems and spikelets of alpine plants注:Ⅰ:矮嵩草;Ⅱ:青藏苔草;Ⅲ:垂穗披碱草。下同Note:Ⅰ:Kobresia humilis;Ⅱ:Carex moorcroftii;Ⅲ:Elymus nutans. The same as below

图3 增温对有性繁殖效力的影响Fig.3 Effect of warming on sexual reproduction efficiency
由图3所示,总体上,矮嵩草有性繁殖效力在增温处理下平均每株减少2%~3%,其变化趋势与繁殖枝数变化趋势基本一致,青藏苔草与垂穗披碱草有性繁殖效力呈下降趋势并与其小穗长度变化趋势基本一致;其中矮嵩草增温下有性繁殖效力极显著降低(P<0.01),而青藏苔草、垂穗披碱草有性繁殖效力无显著变化(P>0.05)。以上结果说明在增温处理下,矮嵩草的繁殖枝数、有性繁殖效力受影响程度较大,与增温呈负响应;青藏苔草、垂穗披碱草的有效穗长度受影响程度较大,且与增温呈正响应。
2.2 种子生产能力的变化
从种子生产能力的角度来看,总体上,增温会引起植物种子生产能力降低(图4)。其中,增温引起矮嵩草植物种子生产能力极显著降低(P<0.01),平均每株矮嵩草所产种子在增温下减少5~6粒;增温引起垂穗披碱草种子生产能力显著降低(P<0.05),平均每株垂穗披碱草所产种子减少13~16粒;青藏苔草种子产量没有显著差异。
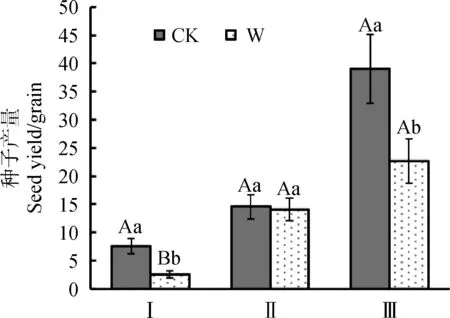
图4 增温对植物种子产量的影响Fig.4 Effect of warming on seed yield
2.3 发芽势与发芽率的变化
在增温处理下,3种植物种子室内发芽势都降低(图5)。其中矮嵩草、垂穗披碱草室内发芽势在增温下极显著降低(P<0.01)。矮嵩草种子发芽势从63.5%降低至22.0%,垂穗披碱草种子发芽势从25.1%降低至6.8%,青藏苔草室内发芽势在增温下显著降低(P<0.05),从6.9%降低至3.6%。3种植物种子室内发芽率在增温下都无显著变化。该结果说明增温处理会降低调查植物种子发芽势,使发芽速度降低,但对其发芽率无明显影响。
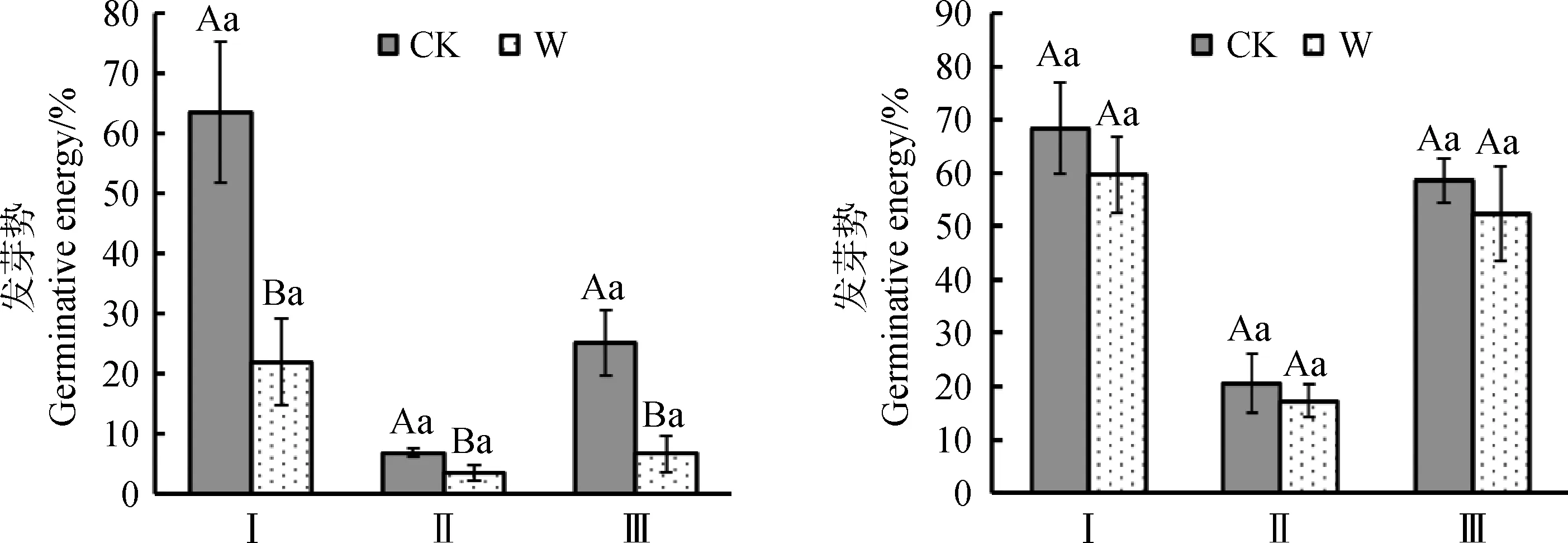
图5 增温对发芽势与发芽率的影响Fig.5 Effect of warming on germination energy and germination rate
2.4 3种植物有性繁殖对增温的响应敏感性分析
根据3种植物有性繁殖指标对增温的响应进行Heatmap作图分析。敏感程度用一定颜色梯度表示(图6中左侧为颜色梯度代表数值),颜色越深代表在增温下该植物的指标越敏感,表示该植物的指标相较其他两种植物对增温的响应程度越大,用可视化的图形综合反应2.1~2.3的结果。
由图6可知,增温下,矮嵩草繁殖枝数、有性繁殖力、发芽势和发芽率指标响应敏感,且幅度较大,青藏苔草的发芽势与发芽率响应敏感;垂穗披碱草与矮嵩草和青藏苔草相比,其小穗长度与种子产量响应敏感,其余不明显。综上,在增温影响下各指标对增温效应响应敏感性最大的是矮嵩草,响应敏感性较大的为青藏苔草,响应敏感性最弱的为垂穗披碱草。

图6 植物有性繁殖对温度响应的敏感性Fig.6 Sensitivity of the response of plant sexual reproduction to temperature注:NBB:繁殖枝数;SRE:有性繁殖力;GE:发芽势;GR:发芽率;SL:小穗长度;SY:种子产量Note:NBB:number of breeding branches;SRE:sexual reproduction efficiency;GE:germination energy;GR:germination rate;SL:spikelet length;SY:seed yield
3 讨论
3.1 增温对3种高寒植物有性繁殖的影响分析
环境因子对植物的有性繁殖特征影响强烈,因此在不同环境因子作用下,草地植物会有很大的形态特征改变甚至会影响植被组成[24]。本研究中利用OTCs气室模拟长期增温探究对3种调查植物有性繁殖的影响,结果表明,增温会使其生殖枝减少而使小穗长度增加,这与王志保等[25]的研究结果一致。由于研究对象的物侯期同样受温度影响[26],长期观测与增温试验发现:随着持续增温,植物返青期或初花期显著提前,秋季枯黄期显著延迟[26],物侯期的变动影响收取种子时间的准确性,若按照原有果实期来进行种子的取样,可能会造成对种子产量、发芽率、发芽势的低估,从而影响试验结果的正确性。但Jiang等[28]在青藏高原研究发现,与其他物候期相比,果期物候对增温和降温的响应保持相对稳定性,这可能是植物维持种子成熟以及传播成功的机制,基于此结论可以确保野外调查时间的准确性。本研究发现,尽管青藏苔草和垂穗披碱草的小穗长度在增温下增加,但种子产量降低,这说明尽管小花数量变多但是结实率反而降低,因此种子产量降低。从结实率低的原因来考虑,由于禾本科和莎草科植物大部分都是风媒花,需要借助风力进行传粉完成受精,且OTCs的“花房效应”限制了水平方向的空气流动,在一定程度上降低了垂直方向的对流过程,室内空气湍流作用减弱[29],极有可能影响了低矮植物矮嵩草、青藏苔草的传粉受精,但垂穗披碱草长势高,可能不受OTCs气室遮挡的影响,能保证传粉自由进行,但依然表现出增温后种子产量降低的效应,这说明增温的确会使植物结实率降低,从而导致其种子产量减少。
植物的有性繁殖阶段容易受到气候变暖的影响[30],且气候变暖可以改变植物的有性繁殖物候,从而大大降低密度依赖和某些植物物种的有性繁殖能力[31],这能很好地解释本研究中矮嵩草在增温处理下各项指标都降低的现象,表明有性繁殖能力被抑制。但也有一些研究表明增温促进植物有性繁殖[29],原因可能是由于增温时间的不同或繁殖对策不同。本研究中,增温后3种植物种子室内发芽势降低,原因可能是在形成种子的过程中,由于增温的影响,加快了土壤水分蒸发速率,植物受到干旱胁迫,在不良环境下易形成休眠种子以渡过不良环境,有利于其种质的延续[32],说明发芽势的降低可能是植物维持种子活力的一种繁殖对策。赵玉红等[33]认为发芽率无明显变化说明种子发芽能力未受损,这就很好解释本研究中增温处理未使种子发芽率明显降低的现象,揭示增温下植物维持种子发芽能力的对策。
综上所述,温度升高一方面将影响土壤含水量及结构,另一方面将影响植物对养分与水分的吸收以及有性繁殖[34],因此推测短期增温促进植物的有性繁殖,但长期增温抑制植物有性繁殖,其原因可能是土壤类型发生改变[29],进而影响高寒草甸植被群落组成和结构,影响群落变化的方向,根据“生草土形成理论”可预测最后会发生植被的演替[32]。
3.2 3种植物有性繁殖对增温的响应敏感性分析
最新数据显示:温度升高使植物有性繁殖过程对气候变化的敏感性增强[30]。不同植物对于温度的敏感程度不同[35]。赵建中等[36]认为不同功能群植物种对地表温度升高的响应是不一致的,地表温度持续升高对禾本科功能群植物种的生长有利,而对莎草科、杂类草功能群物种均不利。这与庞晓瑜[37]研究结论一致,即增温促进禾本科生长,抑制莎草科生长,这能很好解释本研究中矮嵩草的有性繁殖对温度的变化最为敏感,且基本上为负响应,而青藏苔草有性繁殖对温度的响应程度较高,垂穗披碱草有性繁殖对增温的响应程度最低的现象(图6),且对莎草科物种来说不同植物响应程度也是不一致的。从群落中种群的空间分布来考虑,垂穗披碱草处于群落上层,对底层莎草有遮蔽作用,极有可能造成不同功能群之间有性繁殖特征的差异,但是同属莎草科且长势低的矮嵩草与青藏苔草的有性繁殖对温度响应程度也各有不同,因此可以排除垂穗披碱草的遮蔽作用带来的影响,把原因归结为各植物间繁殖对策的差异,使得有性繁殖对温度的响应各有不同,这也就能解释为何增温后青藏苔草与垂穗披碱草的有性繁殖效力无显著变化(图3)。垂穗披碱草的有性繁殖增温下并未明显表示出正响应,因此,若要探究垂穗披碱草是否会取缔矮嵩草成为优势种还需结合无性繁殖进行系统分析。
4 结论
在增温处理下,不同植物表现出不同的繁殖策略,矮嵩草的繁殖枝数量、有性繁殖效力极显著降低(P<0.01)。青藏苔草表现为有效穗长度显著增加和种子产量显著降低的趋势,垂穗披碱草小穗长度极显著增加,种子产量显著降低;增温使3种植物的发芽势有一定下降的趋势,对发芽率影响不明显。在增温下,矮嵩草有性繁殖受到抑制,且最为敏感。
