模拟增温对高寒草甸植物物种多样性与初级生产力的影响
2019-05-31姜风岩位晓婷康濒月邵新庆
姜风岩,位晓婷,康濒月,邵新庆
(中国农业大学草业科学与技术学院草业科学系,北京,100193)
据联合国政府间气候变化专门委员会(Intergovernmental Panelon Climate Change,IPCC)报道,由温室气体排放引起的全球气候变暖最为明显,到21世纪末,地表温度将升高2℃~4℃,上升速度远超过去两千年的任何阶段[1],Thomas等[2]认为在高纬度和高海拔地区温度升幅会更大。气候变暖和降雨格局变化,对生物圈、区域环境、各类生态系统、动植物群落及种群,乃至植物个体生理生态均会产生深刻影响;温度升高直接影响植物体内各种酶的活性、次级代谢产物的合成以及同化物的运输[3],从而影响植物的光合作用和呼吸作用等功能。同时,温度改变土壤营养物质组成和含水量,间接调控着植物的生长[4-5]和生物量生产及分配[6-8],从而引起群落稳定性、初级生产力和生物多样性的变化。气候变化引起的这种改变在高纬度和高海拔生态系统中表现更为明显[9]。
草地初级生产力是草地健康状况的重要指标,是草地生态系统结构和功能的综合体现,是植物生物学特性和外界环境条件共同作用的产物,也是发展牧区畜牧业的基础[10-11]。可见,生产力是一个重要的生态系统功能特征。有研究表明,生产力受生物多样性影响[12],生产力与生物多样性之间的关系因此受到广大学者的关注。
青藏高原由于其严酷而脆弱的环境条件,对气候变化的响应极其敏感[13],被誉为“全球气候变化的驱动机与放大器”[14],是“全球变化与地球系统科学统一研究的最佳天然实验室”[15]。近些年来,研究者就高寒草地多样性及生产力对气候变化的响应开展了一系列的研究工作,但尚未取得一致的研究结论[16-17]。出现不同的原因,一是可能由于植物对气候的响应不仅与地域有关,时空差异、方法差异对结果产生影响;二是因为增温条件下,在未剔除物种的自然群落中的研究较少,所提供的信息尚不足总结增温下高寒草甸群落的响应规律。在当前各研究中,关于增温对高寒针茅化草甸多样性和生产力对气候变化做出何种应答还不明确。本研究基于长期OTCs模拟增温试验,研究高寒针茅化草甸生态系统植物群落物种多样性和初级生产力的变化规律,探讨高寒针茅化草甸对增温效应的响应,为预测全球气候变化对高寒草甸植被的影响以及当地畜牧业的可持续发展提供科学依据。
1 材料与方法
1.1 研究地概况
试验地位于青海省海北藏族自治州海晏县西海镇的海北高原现代生态畜牧业科技示范试验示范园,地处青藏高原东北部青海湖东岸(36 ° 55 ' N,100 ° 57 ' E,海拔3029 m),属于高原大陆性季风气候,年均温1.5℃左右,年降雨量约580 mm,降雨主要集中在7-9月。土壤类型为高山草甸土,土层较浅,植被类型为高寒针茅化草甸草植被,植被总盖度为80%,建群种为紫花针茅(Stipapurpurea),草地早熟禾(Poapratensis),冷地早熟禾(Poacrymophila),披碱草(Elymusdahuricus),线叶嵩草(Kobresiacapillifolia)等,草地类型为高寒草甸草原。
1.2 样地设置
样地始建于2014年6月,样地内设16个小区,设计增温(W,Warming)、对照(CK,Control check)2个处理,每个处理重复8次,采用4行4列完全随机试验设计。每个小区为直径2.2 m圆形,小区与小区之间间隔3 m。增温采用圆底式开顶箱增温装置(OTCs)进行增温2℃,OTCs增温的原理是:由于温室的阻挡作用,OTCs室内风速降低,空气湍流减弱,使热量不易散失,室内温度得以升高[18]。在本试验中,温室内设计了自动控温系统和抽气式风扇,当OTCs内外空气温度差值超过2℃时,风扇自动启动,保证增温不超过设定值。自2014年6月至2018年8月,始终进行全天全年式增温。OTCs永久性安装于样地内,规格为:底部直径2.2 m,顶部直径1.5 m,高度0.7 m(图1)。在16个小区内分别设置一个25 cm×25 cm的样方。
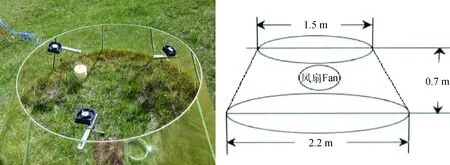
图1 圆底式开顶箱增温装置(OTCs)实景图与示意图Fig.1 On-site picture and schematic diagram of open-top chambers (OTCs)
1.3 测定指标与方法
1.3.1植物调查与物种多样性的计算 于 2018 年 8月中旬取样。用样方法分别在对照样地和OTCs内,进行群落调查和地上生物量的测定,样方面积为25 cm × 25 cm。指标具体测定与计算方法如下[19]:
多度测定:将样方框用细线均分为100个小格,严格统计记录每个物种的个数。
株高测定:在样方内每种植株随机选取10株,用钢卷尺测量从地面到植株自然高度的最高点,求平均值。不足10株的稀缺植株则测定全部株数,并按全部株数求平均值。
盖度测定:将样方框用细线均分为100个小格,通过目测法,对各物种盖度及物种总盖度进行测定。
植物生物量测定:将所有物种齐地面刈割,按照禾本科、豆科、莎草科和其他分类,并分别装入自封袋中带回实验室,称量植物鲜重,记录数据后,在65℃恒温干燥箱干燥48 h至恒重,将干燥后的植物样本称重,得到样方内所有植物种的地上生物量数据。
多样性指数:植被群落多样性指数采用香农-威纳指数(Shannon-Wiener index)、辛普森优势度指数(Simpson dominance index)、均匀度指数(Pielou index)和物种丰富度指数(Species richness index,下文用Q表示)进行分析,其公式[20]如下:
重要值=(相对多度+相对盖度+相对高度+相对频度)/
4×100%
(1)
注:式中相对多度=各物种植株数/所有物种植株数;
相对盖度=各物种的分盖度/物种总盖度;
相对高度=各物种的平均高度/所有物种平均高度之和;
相对频度=各物种在样方中出现的频数/所有物种在样方中出现的频数之和。
Shannon-Wiener指数:
H=-∑PilnPi
(2)
式中Pi=Ni/N;Ni其中为样方中第i种植物的重要值;N为样方植物重要值总和。
Simpson优势度指数:
(3)
均匀度指数:
E=H/lnS
(4)
物种丰富度指数Q:
Q=S
(5)
式中H为Shannon-Wiener指数;S为植物物种数。
1.3.2土壤含水量测定 采用土钻法取样,每个小区随机选取3点,取10~20 cm土层的土壤,将3个点混合为一个土样,分装在3个已称过重的空铝盒中,做好标记,4℃保存。将样品带回实验室称重,在恒温箱65℃下烘干至恒重后再次称重,计算土壤含水量。
土壤含水量=(土壤鲜重-土壤干重)/土壤干重×100%
1.4 数据处理
用Excel 2013进行数据统计及初步分析,并绘图;SPSS 21.0对各处理物种的重要值、功能群的植被多样性及初级生产力进行t检验,对植被多样性与初级生产力进行相关性分析。
2 结果与分析
2.1 增温对植被群落组成的影响
在禾本科、豆科、莎草科和其他科植物中,分别选择重要值相对较高的物种,对其在增温下重要值的变化进行分析。在禾本科中,选择了5个物种,其中,针茅(Stipacapillata)、克氏针茅(Stipakrylovii)的重要值下降显著(P<0.05),洽草(Koeleriacristata)的重要值则显著升高(P<0.05)(图2)。在豆科与莎草科中,各选择了2个物种。增温下矮嵩草(Kobresiahunilis)、苔草(Carextristachya)的重要值变化显著(P<0.05),但是变化不一致,矮嵩草重要值上升,苔草重要值下降。多枝黄耆(Astragaluspolycladus)与扁蓿豆(Melissitusruthenicus)的重要值无显著变化。其他科植物中,共选择了5个物种。多裂委陵菜(Potentillamultifida)、米蒿(Artemisiadalai-lamae)、猪毛蒿(Artemisiascoparia)的重要值在增温处理下显著下降(P<0.05),鳞叶龙胆(Gentianasquarrosa)重要值在增温下显著上升(P<0.05),异叶青兰(Dracocephalumheterophyllum)重要值无显著变化(图2)。总的来看,增温处理下,群落的物种组成发生了显著的变化,矮嵩草重要值增加而针茅重要值下降,针茅不再具有绝对优势,但是洽草的重要值显著上升。可见,禾本科植物在群落中仍然占据很大比例。
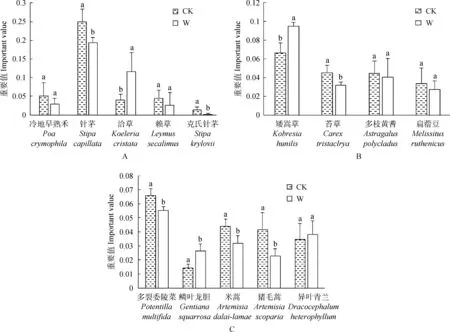
图2 增温对群落物种重要值的影响Fig.2 Effect of warming on important values of community species注:A为禾本科,B为莎草科和豆科,C为其他类;每个图柱上方不同小写字母表示在0.05水平上差异,下同Note:A is Graminease,B is Cyperaceae and Leguminosae,and C is others;Different lowercase letters above each column indicate differences at the 0.05 level,the same as below
2.2 增温对植被群落多样性的影响
研究结果显示,增温使高寒草甸Shannon-Wiener多样性指数显著下降(P<0.05)。Pielou均匀度指数略有升高,与对照相比变化不显著。虽然群落中一些物种的重要值发生了显著性的变化使得Simpson优势度指数有所下降,但变化不显著。物种的丰富度指数显著降低(P<0.05)。(图3)
2.3 增温对初级生产力及土壤含水量的影响
与对照相比,在增温处理下,禾本科、莎草科、其他科植物的初级生产力均有所降低(图4)。其中,其他科植物生产力的变化达到显著水平(P<0.05)。增温条件下,豆科的初级生产力有所增加,但差异不显著(图4)。增温处理下群落总生产力降低但差异不显著。就生物量在各科的分配比例而言,禾本科占比最大,在对照中占比达到65.6%,在增温下达到64.6%;莎草科生物量在对照和增温中占比分别为15.8%和11.6%;豆科生物量在对照和增温中占比分别为4.6%和14.5%。增温处理下,土壤含水量显著下降(P<0.05)(图5)。
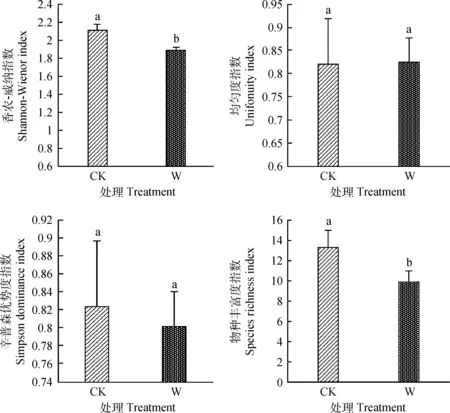
图3 增温对物种多样性的影响Fig.3 Effect of warming on species diversity characterized by three indices
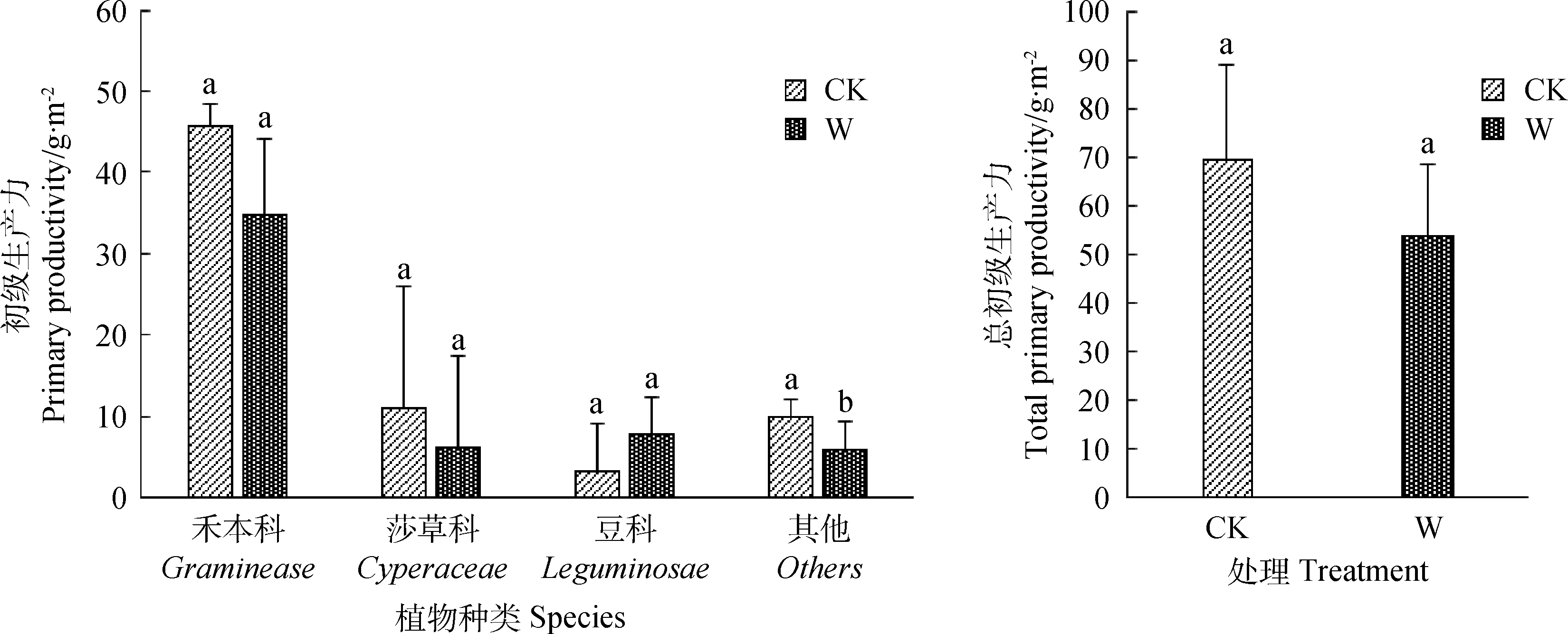
图4 增温对各科及总初级生产力的影响Fig.4 Effect of warming on various subjects and total primary productivity
2.4 物种多样性与初级生产力相关性分析
高寒草原植物物种多样性与初级生产力之间的拟合关系和相关性系数见表1。拟合结果显示,在对照中,丰富度指数与初级生产力的相关性较差,均匀度指数与Shannon-Wiener指数与初级生产力的相关性较高,相关系数R2均在0.65以上。增温处理下,均匀度指数和Shannon-Wiener指数与初级生产力的相关性仍大于丰富度指数,相关系数R2均大于0.43。增温降低了物种多样性与植被生产力间的相关性。
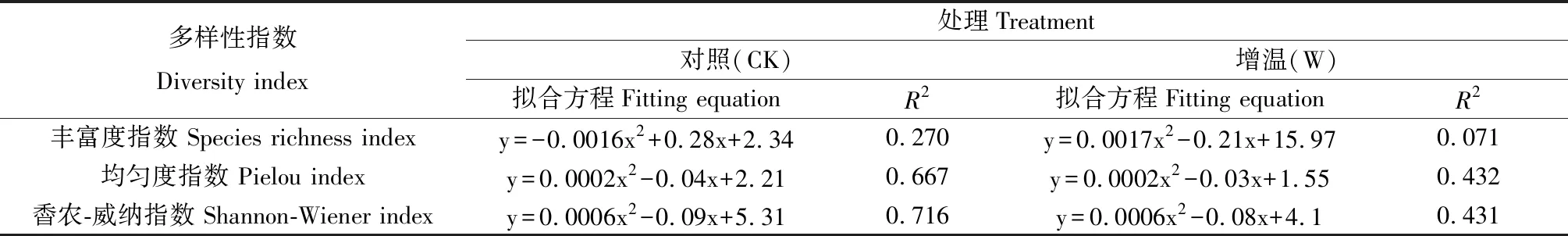
表1 不同处理下物种多样性指数与初级生产力的关系拟合Table 1 Relationship between plant species diversity index and primary productivity under different treatments
3 讨论
3.1 增温对群落结构及初级生产力的影响
温度能控制生态系统中许多生物化学反应速率,且几乎影响着所有生物学过程[21]。温度升高在一定程度上能够满足植物对热量的需求,但也改变了植物群落的小气候环境,从而通过多种途径直接或间接影响植物的生长发育及生物量生产[22]。不同持续时间的增温对植物群落的影响不同。Zhang等[23]和周华坤等[24]通过1年的增温试验得出,在短期增温条件下群落的初级生产力变化不明显;李英年等[25]通过5年的增温试验研究表明,在模拟增温初期初级生产力增加,但增温5年后初级生产力反而下降。在本研究中,连续4年的增温处理下,仅其他类草的初级生产力出现显著下降。可见,增温对群落初级生产力影响的正负效应是一种随时间而持续变化的过程,而这个变化的结果很可能与土壤的含水量相关。
本试验结果表明,与对照相比,土壤含水量显著降低,这与当前多数试验结果一致[26-27]。OTCs装置改变了植物群落生长的小气候,使小室内温度长期处于升高状态,这直接加快了土壤中水分的蒸散,使小室内的环境逐渐趋向暖干化。而对于青藏高原高寒草原的植物来说,低温和水分是限制高寒草地植物生长的两个关键因素[24]。因此,环境的暖干化,是诱发生态变异的动力[28]。短期增温处理,满足了高寒植物对温度的需要,使得植物体内各种化学反应的速率加快,此时水分的限制条件也不明显[28],初级生产力因此上升。而长期的增温导致表层土壤长期处于缺水干旱状态,此时的水分成为植物生长的限制因素[3]。可以认为,水分胁迫是短期与长期增温条件下,高寒草甸初级生产力的变化有明显差异的原因之一。
增温影响了植物对水分的吸收,进而影响植物生物量的积累[24,29-30],而不同种植物积累生物量的变化程度也不相同。大量的研究结果表明[5-6,28],在全球变暖背景下,对于任何一个植物群落来说,总有一些物种比另一些物种对温度升高的响应更为敏感,从而破坏种间竞争关系,引起群落的优势种和组成发生改变。本研究结果显示,增温显著降低了其他类草的生物量(P<0.05),禾本科与莎草科植物的生物量也有下降趋势,这说明增温引起的水分胁迫对高寒草原群落的影响较大,大多数植物选择减少地上生物量以应对土壤水分的降低。
增温对各物种生长的影响不尽相同。有研究显示,增温后表层土壤缺水干旱,严重限制了植物根系在表层的生长[31]。植物根系受到影响,自身的生长会发生改变。在本研究中,增温使得针茅、克氏针茅、苔草、多裂委陵菜、米蒿、猪毛蒿的重要值显著下降,这说明对于上述植物,土壤水分降低带来的胁迫可能超过了温度升高的促进作用。冷地早熟禾、赖草、多枝黄耆、扁蓿豆以及异叶青兰的重要值变化不显著,说明在一定的增温促进与水分胁迫的影响下,这些植物仍能较好的适应。莎草一般较禾草耐阴,当禾草植物占据群落上层时,形成郁闭的环境,莎草科成为群落的下层[24]。在增温的影响下,禾草的生长受到一定抑制,矮嵩草可能因此获得更多的光照资源,重要值得到显著提升(P<0.05)。而同为莎草科的苔草则可能因为与矮嵩草产生竞争导致重要值显著下降(P<0.05)。可见,不同物种对增温以及增温造成的水分胁迫的响应不同。
不同物种对温度响应的差异使原有群落结构的稳定性被打破,群落的组成发生改变。本研究中,增温降低了群落物种数,但是能够适应环境变化的物种相对密度有所增加,达到一种新的群落结构的稳定,所以植物群落的均匀度指数没有显著性变化。Shannon-Wiener指数不仅与植物的相对密度有关,也与植物的物种数有关,所以,增温处理下,Shannon-Wiener指数显著下降(P<0.05)。
3.2 植物多样性与初级生产力的关系
本研究中,物种多样性与初级生产力的最佳拟合模型是二次多项式。试验结果也表明,在增温处理下,Shannon-Wiener指数显著下降(P<0.05),而群落的初级生产力没有发生显著变化,这说明群落的初级生产力与物种多样性不是简单的线性关系。在自然生态系统中,物种多样性和生产力受到非生物因素(资源)的影响,而这些非生物因素有可能是群落多样性和生产力关系的幕后操纵者[32]。有研究发现,光照和土壤肥力会同时影响植物多样性与生产力的关系[33]。本研究发现增温处理下物种多样性与初级生产力的相关系数降低,说明增温使物种多样性与初级生产力的相关性减弱。
生态位互补效应表明,与单一的功能群相比,功能群数目的增加会使植物对资源的互补利用程度增加,资源利用效率提高,从而提高群落生产力[34]。资源的匮乏会使资源的利用受到限制,可能使得物种间不能体现出对资源的充分利用能力,即多样性的作用缺乏表现条件[33],进而使两者之间的关系减弱。此外,不同的功能群划分方法也可能会对多样性和生产力间的非线性关系产生影响[35]。
物种多样性与初级生产力的非线性关系说明仅以物种多样性来研究群落的初级生产力是不全面的,董世魁等[36]的研究结果表明群落的初级生产力还会受到功能多样性的影响。因此,在研究高寒草甸群落的初级生产力与多样性的关系时,不能只考虑物种多样性这一种方面,还应考虑功能多样性,以期更准确地表述高寒草甸的生态功能状态。
4 结论
综上所述,增温使得青藏高原高寒草甸的土壤相对含水量显著下降(P<0.05),而水分是限制该地区植物生长的关键因素之一。增温使得高寒草甸的物种重要值和群落组成发生改变。群落的物种丰富度显著下降(P<0.05),Shannon-Wiener指数显著下降(P<0.05)。群落总的初级生产力变化不显著,但群落中其他类植物的生产力显著下降(P<0.05)。各种类型植物的生产力占比有所改变,这很可能会对家畜营养需求产生一定的影响。物种多样性与初级生产力之间不是简单的线性关系,增温引起的水资源匮乏会导致两者的相关性减弱。植物多样性与初级生产力的关系会受到诸多环境因素的影响,因此进行研究时,要综合考虑物种多样性与功能多样性。
