四川华鳊精子发生显微及超微结构观察
2019-04-08解崇友王志坚
尹 敏,解崇友,金 丽,王志坚
(西南大学生命科学学院,淡水鱼类资源与生殖发育教育部重点实验室,水产科学重庆市重点实验室,重庆 400715)
四川华鳊(Sinibramataeniatus)属鲤形目(Cypriniformes)鲤科(Cyprinidae)鲌亚科(Cultrinae)华鯿属(Sinibrama),仅分布于长江上游岷江、青衣江、嘉陵江以及大渡河等水域[1],是长江上游珍稀特有小型鱼类。近年来,长江上游干流及主要支流大规模修建电站,拦河筑坝,致其生境尤其是繁殖环境遭到不同程度破坏,持续的高强度捕捞压力也使得四川华鳊种群数量锐减,解崇友等[2]的研究发现,四川华鳊的种群已出现小型化趋势,因此有必要开展四川华鳊种质资源保护相关工作。目前关于四川华鳊的研究资料主要集中在资源调查[3]、年龄生长[2]以及繁殖生物学[4]等,未见室内人工繁殖的相关研究。人工繁殖是实现渔业资源保护和自然种群恢复、增殖的重要技术手段,而生殖细胞发生作为繁殖生物学的重要组成部分,了解其在生殖周期中的动态变化,对于掌握物种繁殖规律具有重要意义。本研究以四川华鳊成鱼为研究对象,通过性腺组织学切片,辅以透射电镜技术,对其精子发生过程进行显微及超微结构的观察,丰富其繁殖生物学基础资料,为人工繁殖提供科学指导。
1 材料与方法
1.1 实验材料
岷江眉山段采集的性成熟四川华鳊,在实验室条件下暂养两周后开始取材,每月一次。养殖条件:自然光照,水温(20±2) ℃,pH值7.5±0.5,溶解氧>7 mg/L,每日9∶00投喂浮性颗粒饲料。
1.2 实验方法
解剖取四川华鳊成鱼精巢,显微观察采用Bouin氏液固定24 h,梯度酒精脱水,二甲苯透明,石蜡包埋,Leica RM 2235切片机切片5 μm,H.E染色[5],Nikon Eclipse 80 i显微镜观察并拍照;超微结构观察采用预冷的4%戊二醛固定,经0.1 mol/L磷酸缓冲液(pH值7.4)冲洗3次,1%锇酸后固定1~2 h,双蒸水清洗3次(每次不超过3 min),梯度酒精脱水(50%乙醇,15 min;70%乙醇+醋酸双氧铀过饱和溶液,过夜;90%乙醇,10 min),丙酮置换(90%乙醇+90%丙酮1∶1,10 min;90%丙酮,10 min;100%丙酮,10 min,3次),环氧树脂650聚合包埋,LBK-5型超薄切片机切片,醋酸超氧铀和枸櫞酸铅双染,JEM-1400 Plus透射电镜观察并拍照。
1.3 数据处理
使用图像分析软件Image-Pro Plus 6.0测量生殖细胞直径及核径,SPSS 17.0对数据进行统计分析。
2 结果
2.1 生殖细胞形态学指标
光学显微镜及透射电镜下随机选取不同标本的不同区域进行拍照,统计结果见表1。

表1 四川华鳊雄性生殖细胞大小统计表Tab.1 Statistics of male germ cell sizes in S. taeniatus μm
2.2 生殖细胞发生的显微及超微结构
2.2.1 精原细胞
精原细胞在成鱼性腺的各个时期中都能找到,精原细胞沿精小叶分布,一般在精巢边缘分布较多(图版I-1)。精原细胞为精子发生过程中体积最大的细胞,H.E染色后,精原细胞着色最浅,可分为两种类型,分别称A型精原细胞和B型精原细胞。A型精原细胞为精原干细胞,细胞卵圆形,细胞长径(9.80±0.62) μm,细胞核也为卵圆形,长径(6.28±0.50) μm,核膜清晰,核仁明显,位于细胞核中央或偏向一侧,嗜碱性强,染成蓝色。核质中有极少的嗜碱性物质也着蓝色,细胞质与细胞膜均不着色,因此细胞边界不清晰但高倍镜下基本能分清细胞界限(图版I-2)。A型精原细胞分裂产生B型精原细胞。B型精原细胞稍大,长椭圆形,细胞直径的长径为(11.02±1.13) μm,细胞核长径为(7.42±0.82) μm,光学显微镜下B型精原细胞核仁不明显,核内嗜碱性染色质分散于核膜边缘或核质内各处(图版I-2)。
两种类型的精原细胞在显微结构上只能根据核仁大小及核膜是否清晰加以区分,而超微结构显示,二者除了在核仁大小、电子密度以及核膜上的区别外,其胞质中拟染色质小体的数量、线粒体的多少等也是区分两种细胞的重要依据。A型精原细胞的细胞膜及核膜清晰,中央大核仁电子密度高,核内染色质均一,核膜外周有多个拟染色质小体,核内有与其电子密度相似的颗粒物质,胞质中线粒体丰富,內嵴清晰,透射电镜下整体呈棒状或椭圆形(图版II-7)。B型精原细胞其核膜较前者模糊,核仁松散,电子密度较低,核质均一,核膜外周拟染色质小体较少,胞质中线粒体明显减少。两种类型的精原细胞在与相邻细胞接触的位置均出现了细胞膜的缺口,猜测为细胞质桥(图版II-8)。
2.2.2 初级精母细胞
B型精原细胞经有丝分裂生长形成初级精母细胞,初级精母细胞主要存在于Ⅱ期和Ⅲ期精巢中,在较晚时期的精巢中数量不多。初级精母细胞长径(8.08±0.46) μm,细胞核长径(4.95±0.58) μm,光学显微镜下只能观察到初级精母细胞的细胞核,细胞边界几乎不可见,与精原细胞相比,细胞核质内染色质凝缩,丝状或团块状不均一分布,胞核嗜碱性增强,H.E染色后呈浅蓝色,核膜清晰,细胞核形态不一,部分初级精母细胞有中央大核仁,呈深蓝色,部分初级精母细胞无明显的中央大核仁(图版I-3)。
超微结构中观察到的初级精母细胞为不规则长椭形,细胞核圆形,胞质中细胞器较多,核质比增大不明显,仍能观察到核膜的双层膜结构,核膜开始出现不同程度波曲,核质电子密度较精原细胞大,核内染色质团块状不均匀分布,核膜外有拟染色质小体存在,膨大线粒体附着在拟染色质小体附近,胞质中也有许多线粒体分布,可观察到内质网,未观察到具中央大核仁的初级精母细胞,在初级精母细胞群附近观察到了桥粒连接(图版II-9)。
2.2.3 次级精母细胞
初级精母细胞在减数第一次分裂后形成次级精母细胞,次级精母细胞大量存在于III期、IV期及V期早期的精巢中,其体积进一步减小,细胞长径(4.75±0.49) μm,约为初级精母细胞的一半,细胞核长径(3.75±0.24) μm,核质比明显增大,核内染色质进一步凝缩,高倍镜下可观察到核质线团状散乱分布,核质嗜碱性增强,整个细胞核染成较均一的蓝色(图版I-4)。
超微结构中观察到的次级精母细胞呈卵圆形,细胞核形态与初级精母细胞相似,近圆形,细胞膜的双层膜结构不清晰,但可根据电子密度区分细胞核与胞质,核内染色质均一,电子密度增大,核膜附近未见拟染色质小体,胞质内仍分布有大量线粒体,但内嵴不明显,此外,胞质内还有许多电子密度较大的颗粒状物质。同样,次级精母细胞间也有细胞质桥存在(图版II-10)。
2.2.4 精子细胞
次级精母细胞通过减数第二次分裂产生精细胞,在Ⅳ期的精巢中较易观察到精细胞,精细胞长径(3.81±0.38) μm,较次级精母细胞进一步缩小,核长径(2.94±0.36) μm,光学显微镜下观察到精细胞的细胞核圆形,较精母细胞体积明显减小,只比精子稍大,细胞整体呈深蓝色(图版I-5)。

图版I 四川华鳊精子发生的显微结构(H.E 染色)Plate.I Microstructure of Sinibrama taeniatus spermatogenesis
电镜下观察到的精细胞有多种形态,根据核质浓缩程度及细胞形态结构特征,可将精细胞产生到成熟精子形成之前的过程大致分为三个阶段,早期的精细胞体积较次级精母细胞稍小,细胞间开始出现明显缝隙,但仍有少量的细胞质桥连接,核质电子密度稍有增加,核质比增大,胞质减少并向一侧集中,胞质内可见多个膨大的线粒体,高尔基体结构明显,大部分精细胞在精子头部正下方的胞质中有一空腔,内有形似微管的结构,猜测为基体的横切面。在结构完整的早期精细胞中,观察到互相垂直的基体和近端中心粒,已有鞭毛沿基体伸出(图版II-11)。中期的精细胞体积进一步减小,没有多余胞质,精子头部基本形成,细胞边缘可观察到多层膜结构,由细胞膜及核膜紧贴形成,细胞核进一步浓缩,核质密度明显增大,可观察到核空泡,高尔基体和膨大的线粒体位于胞核后端两侧的胞质内,胞核正下方为内含基体结构的腔(图版II-12)。晚期的精细胞中,由基体延伸出的鞭毛仍位于胞质下方的空腔内,未见结构清晰的近端中心粒,但在基体上方靠近核膜的位置有中心粒附属物的形成(图版II-13)。
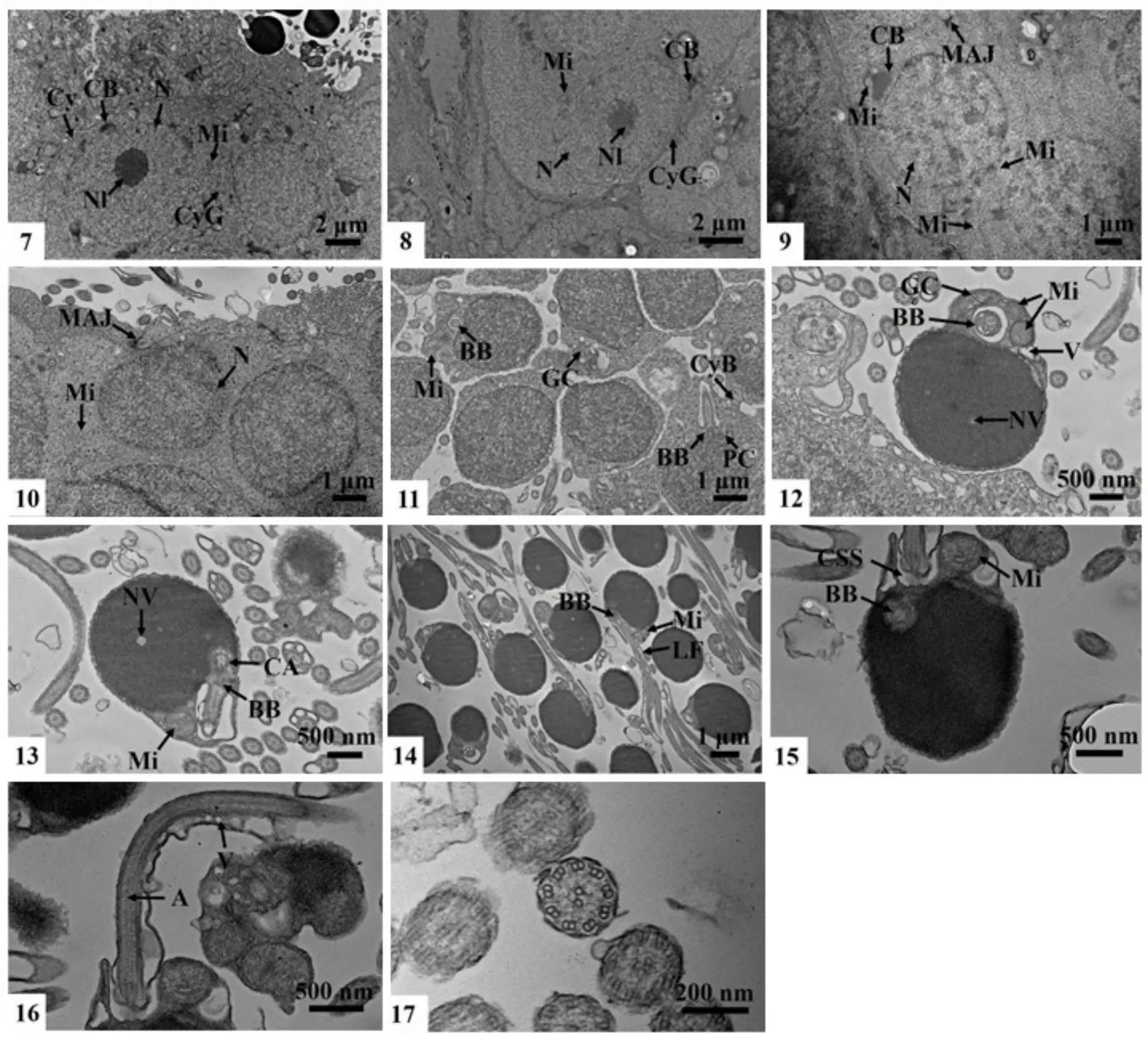
图版II 四川华鳊精子发生的超微结构Plate.II Ultrastructure of Sinibrama taeniatus spermatogenesis
2.2.5 精子
精子细胞经变态发育为成熟精子,精子大量存在于Ⅳ期及Ⅴ期精巢的小叶腔中,在早期的精巢中也可观察到少量的成熟精子。精子嗜碱性极强,为染色最深的生殖细胞,呈深蓝色(图版I-6)。
电镜下观察到的精子分三部分(图版II-14):精子头部近圆形或稍不规则,无顶体,长径(2.42±0.02) μm,核长径(2.28±0.05) μm,绝大部分为细胞核,电子密度极大,电镜下呈黑色,精子头部切面可观察到核空泡,但较小,核凹窝不发达(图版II-15);精子中片紧靠精子头部下端核膜,由精子尾部着生位置的中心粒复合体和袖套共同构成,在成熟精子中,中心粒复合体中最明显的结构为基体,未见中心粒及其附属物,精子尾部着生位置胞质凹陷形成袖套腔,腔内有膨大的线粒体及一些囊泡化的细胞器结构,精子尾部由轴丝构成,贯穿精子细长尾部,形成精子运动的支撑结构,精子鞭毛横切可观察到轴丝由外围9组二联体微管及中央2根独立微管共同组成,精子尾部两侧有侧鳍,基本对称,精子尾部后端的侧鳍中有囊泡存在(图版II-15,16,17)。
3 讨论
四川华鳊精子发生过程经历了精原细胞、初级精母细胞、次级精母细胞以及精子细胞变态的过程。A型精原细胞通过有丝分裂形成B型精原细胞,B型精原细胞不断进行有丝分裂,产生能够进入减数分裂的初级精母细胞,初级精母细胞经两次减数分裂形成精细胞,再经变态形成成熟精子。在这个过程中,除两次减数分裂以外,生精细胞的细胞核及细胞质结构均发生了一系列变化。
精原细胞核中最显著的结构为中央大核仁,但B型精原细胞的核仁更为松散。核仁与蛋白质的旺盛合成、细胞的活跃生长有关[6],核仁一般在分裂前期消失,分裂末期重新出现。在四川华鳊的两种精原细胞中均观察到了核仁,而在显微结构中只有部分初级精母细胞可观察到明显核仁。康现江等[7]发现半滑舌鳎的初级精母细胞核仁在减数第一次分裂前期的偶线期消失,与本研究的结果基本相符,但没有说明核仁是否在减数第一次分裂末期重建,可能由于减数第一次分裂完成后停留时间太短,不易观察到该阶段的生精细胞。四川华鳊精子发生过程中,初级精母细胞之后的生精细胞中也未观察到明显核仁。
在超微结构中,从精原细胞分裂开始,处于同一精小囊内的生精细胞间就出现了细胞质桥,具体表现为相邻细胞在接触位置细胞膜出现了明显缺口,这种现象直至发育到精细胞早期才结束。这与同一精小囊内生精细胞的同步发育有关,这与车静[8]对大眼鳜精子发育的研究结果一致,而管汀鹭等[9]在对金鱼精子的研究中,将这一结构称为细胞间桥,并认为细胞间桥直至精子成熟前才消失。然而,关于细胞质桥的具体定义,不同的研究者持有不同观点。洪万树等[10]在对鲻鱼精子发生的超微结构观察中发现,初级精母细胞有丝分裂过程为不完全胞质分裂,胞质之间出现合胞体,从而导致细胞界限不明显,不同的是,他们仅将精细胞间的连通结构称为细胞间桥。郭申明等[11]也提到,在孔雀鱼的精子发生过程中,从精母细胞到精子细胞其胞质均有不完全分裂现象,但未将这样的结构称为细胞质桥或细胞间桥。不管称为合胞体的胞质连接还是细胞质桥,这一现象的存在都是精小囊内生精细胞维持高效同步发育的前提。大量研究表明,细胞质桥在各种生物精子发生过程中具有普遍性,从低等动物如带绦虫[12]、人皮蝇[13]等到哺乳动物如大鼠[14]以及人的精子发生过程中,生精细胞的分裂过程始终伴随着相邻细胞胞质的沟通。
在四川华鳊精子发生过程中,两种类型的精原细胞以及初级精母细胞的核膜外侧均存在体积较大、数量不等的拟染色质小体,同时在精原细胞核内观察到了与其电子密度相似的颗粒,猜测拟染色质小体的形成与核内的这些高密度颗粒物质有关。根据管汀鹭[15]对金鱼精子发生中拟染色质小体的研究,细胞核内物质聚集形成致密体,排入胞质后形成拟染色质小体,在次级精母细胞以后,核内不再形成致密体,胞质内的拟染色质小体被消耗,数量和体积减小,这可以解释为什么在次级精母细胞及以后的细胞中很难观察到拟染色质小体。Walt等[16]认为拟染色质小体内存在肌动蛋白,与高尔基体共同作用参与顶体蛋白的形成,并参与RNA代谢。
精子变态过程中细胞质偏向一侧,细胞核浓缩,在这一过程中,中心粒复合体起着至关重要的作用。四川华鳊精子细胞变态早期就已出现近端中心粒和基体,变态中期出现了中心粒附属物,而成熟精子中只观察到有基体的存在。根据林丹君等[17]对褐菖鲉精子的描述,中心粒复合体在精子细胞晚期发生变化,中心粒附属物形成,在精子成熟前,中心粒附属物萎缩直至消失,近端中心粒也在精子成熟时消失,这与我们的观察结果基本一致。Fawcett等[18]认为,中心粒附属物可能与哺乳类精子颈段的形成有关,但尚未有研究提到其在硬骨鱼类精子形成过程中的作用。王志坚等[19]对长薄鳅精子发生的研究中,认为远端中心粒(基体)向细胞后部移动导致胞质被拉长,近端中心粒向细胞核方向迁移导致核质进一步浓缩,形成核凹窝。四川华鳊精子的核凹窝极浅,核空泡小,侧面说明其精子变态过程中近端中心粒向细胞核方向迁移程度不大,核质浓缩程度较小,这一特性在其精子头部的形态上也有所体现。四川华鳊精子头部与草鱼[20]、刀鲚[21]、泥鳅[22]等相似,而与长吻鮠[23]、胭脂鱼[24]等核凹窝极深的精子形态不同。研究显示,同为鳅科的泥鳅[22]、大鳞副泥鳅[25]、宽体沙鳅[26]三种鱼类精子头部均为圆形,核凹窝浅,鲿科鮠属的短尾鮠[27]、粗唇鮠[28]的精子头部形态则与长吻鮠相似,核凹窝都较深,张涛等[29]对同属四种罗非鱼精子的对比研究显示,其精子头部结构大体相同,推测精子头部的形态特征可能具有种属特异性,或可作为鱼类分类鉴定的一项指标及精子结构预测的依据,但这一猜想尚待进一步验证。
与绝大多数体外受精鱼类[30-32]精子一样,四川华鳊精子为无顶体型,尾部具两侧对称的侧鳍。侧鳍的有无、发达程度以及形态结构在不同硬骨鱼中具有差异性。目前的研究表明,许多硬骨鱼精子尾部均具侧鳍,为胞质延伸形成。胭脂鱼[24]、黄颡鱼[33]精子尾部侧鳍发达,两侧基本对称,鲻鱼[10]精子尾部侧鳍亦为两侧对称,但发达程度不如四川华鳊精子。江鳕[34]、银鲳[35]精子尾部侧鳍不对称,泥鳅[22]、叉尾鲶[36]精子尾部无侧鳍。关于精子尾部侧鳍的生理意义,不同研究者持有不同意见:Stoss[37]认为侧鳍的存在能提高精子的运动速度,而尹洪滨等[38]通过对不具侧鳍的索氏六须鲶精子的研究后,认为精子运动速度主要取决于鞭毛,侧鳍只起到部分作用。但目前尚未有研究者对不同类型精子做过对比研究,要明确侧鳍的具体作用还需进一步研究。
