茶小绿叶蝉转录组Bt潜在受体分析
2019-01-25陈明峰林桂芳蒋晓燕刘文诚KhadijaBATOOL张灵玲
陈明峰, 林桂芳, 蒋晓燕, 王 瑞, 许 瑾, 刘文诚, Khadija BATOOL, 关 雄, 张灵玲
(福建农林大学生命科学学院,福建 福州 350002)
茶小绿叶蝉(Empoascaflavescens)是茶树上三大害虫之一[1].该虫为同翅目(Homoprtera)昆虫,有虫体小、繁殖能力强、叠代情况严重等特点.该虫有刺吸式口器,以茶树嫩芽叶为食,会造成叶脉、叶片、芽叶等出现不同程度的变红、萎缩甚至掉叶等现象,直接影响茶叶的产量和茶产品的品质.我国江、浙、皖、赣、鄂、湘、蜀、闽、粤、桂等省、区茶叶主产区都受到了茶小绿叶蝉不同程度的危害[2].
苏云金芽孢杆菌(Bacillusthuringiensis,Bt)因为对环境友好、杀虫谱广且对人畜无害而成为当今世界上研究最深入、应用最广泛的一类微生物杀虫剂[3].Bt Cry蛋白是一种成孔细胞毒素,其杀虫活性主要取决于毒蛋白活性片段的酶解及其与特异受体结合后在中肠上皮细胞刷状缘膜蛋白上形成离子通道的能力.目前,已经基本明确的Bt受体有APN[4]、Cadherin_[5]、ALP[6]、肌动蛋白[7]以及glycolipids[8]等.Bt的杀虫机理决定了Bt杀虫蛋白对昆虫中肠受体具有选择性[9];同时,毒素受体缺乏或突变会使昆虫与杀虫蛋白结合能力下降或丧失,或者导致昆虫对Bt产生耐药性.研究表明,Bt对茶小绿叶蝉无明显杀虫效果,有关其机制尚不明确[10].课题组前期已对该虫进行了转录组测序分析,并开展了茶小绿叶蝉肠液对部分Cry毒素的酶解分析,发现虽然该虫肠液对Cry1Ac和Cry11A酶解程度不够彻底,但仍有相当比例的毒素可被酶解成活性片段[11].因此,本研究从Bt杀虫的另一个关键步骤——受体与毒素活性片段的结合着手,依据实验室前期获得的茶小绿叶蝉转录组数据,对该虫的Bt潜在受体基因进行分析,以期为进一步明确Bt对茶小绿叶蝉的作用机制提供一定理论依据,也为开发新型生物农药奠定理论基础.
1 材料与方法
1.1 供试昆虫
本研究所用茶小绿叶蝉采集于福建省福安市农业科学院茶叶研究所2号山平地品种为金观音的茶树.
1.2 试剂
E.Z.N.A. HP Total RNA Kit由OMEGA提供;反转录试剂盒购自TOYOBO公司;250 bp DNA Ladder Marker购自TaKaRa公司;荧光定量试剂盒购于Roche Company;溴化乙锭染色液购于TaKaRa公司.TAE(50×)配制:称取242 g Tris-base、37.2 g Na2EDTA·2H2O,先用适量的灭菌水溶解,定容至1 L,再用57.1 mL乙酸搅拌至完全溶解.
1.3 方法
1.3.1 茶小绿叶蝉转录组Bt潜在受体基因的筛选 根据课题组前期对茶小绿叶蝉转录组RNA-seq得到的Unigene注释了transcripts(NCBI BioProject ID: SRP075995)[12].利用这些数据并依据关键字(ALP、APN、Cadherin)和GPI瞄定位点信息,从de novo组装注释的转录本找出注释为ALP、APN、Cadherin的功能基因;利用RNA-seq得到差异性表达数据库中的表达量,即FPKM值(每百万Reads中来自比对到某一基因每千碱基长度的Reads数目),直接比较不同样品间3类潜在受体基因的表达差异.以FPKM值为指标,将FDR(false discovery rate)<0.01作为阈值,且差异倍数FC(fold change)≥2作为筛选标准(其中FC表示2个样品表达量的比值),筛选出可表达的3类潜在受体基因,并比较其在茶小绿叶蝉中肠和肠外组织中的表达量,从而筛选出茶小绿叶蝉Bt Cry潜在受体靶基因.
1.3.2 Heatmap作图分析潜在受体基因 利用在de novo组装注释的转录本找到注释为ALP、APN、Cadherin功能的基因,再根据差异性分析中计算的表达量进行FPKM值分析,找出候选功能基因在茶小绿叶蝉中肠和肠外组织的表达量.用R语言Heatmap软件作图,通过热点分布情况对茶小绿叶蝉肠道及肠外组织ALP、APN、Cadherin基因的相对表达量进行可视化分析,以找出在中肠中表达量高且与肠外组织表达量差异显著的潜在受体基因.
1.3.3 RT-qPCR验证 参照OMEGA总RNA提取试剂盒操作说明提取茶小绿叶蝉总RNA,并从转录组数据中筛选出8个ALP、7个APN和8个Cadherin差异表达候选基因,设计并委托铂尚生物公司合成引物,以茶小绿叶蝉actin基因(GenBank登录号KJ942579.1)为内参基因[13].利用TOYOBO反转录试剂盒将总RNA反转录成cDNA,然后利用荧光定量试剂盒在ABI7500实时荧光定量PCR检测系统上进行RT-PCR反应[11].
每个样品的实时荧光定量PCR运行完成后进行溶解曲线分析,以检查PCR反应的特异性.具体操作步骤:根据RT-qPCR得到的数据,利用溶解曲线分析计算2个组织中所获得的相对表达量数值,相对表达量计算方法为虫尸表达量/肠外组织表达量;同时,利用Excel做出RT-qPCR得到的各基因的相对表达量和RNA-seq得到的FPKM值这2组数据的散点图.从表达相关性系数R2验证RNA-seq的准确性,R2值越接近于1表示2种方法计算的表达量越相似,说明RNA-seq的结果就越可靠.
1.3.4 受体蛋白氨基酸序列同源性分析 利用关键字(ALP、APN、Cadherin)搜索在茶小绿叶蝉de novo组装注释的转录本,寻找肠内外表达差异显著的ALP、APN、Cadherin 3类23个潜在受体蛋白,在转录本中找到各自对应的氨基酸序列.借助Mega(Version 7.0)软件构建NJ进化树,对ALP、APN、Cadherin 3类潜在受体表达量差异显著的基因序列进行同源性分析.
2 结果与分析
2.1 茶小绿叶蝉潜在受体筛选
由于目前未获得茶小绿叶蝉的基因组测序信息,以NCBI非冗余(non-redundant,nr)数据库和Swiss-Prot数据库作为参考,根据鳞翅目、双翅目、半翅目等昆虫受体信息,选择APN、ALP、Cadherin为茶小绿叶蝉Cry蛋白候选潜在受体类型.通过BLAST进行transcripts的注释,结果获得了52 182个组装的Unigene.利用关键字筛选转录组测序数据中Bt在茶小绿叶蝉中肠和肠外组织中差异表达的潜在受体基因,发现共有3 327个,其中1 191个上调表达,2 136个下调表达.
按照FC≥2、FDR<0.01的原则,筛选获得123个显著差异表达基因,包括13个APN、25个ALP、85个Cadherin.其中,在中肠中表达量高且与肠外表达量差异显著的APN、ALP、Cadherin基因有23个,依序分别重命名(表1).
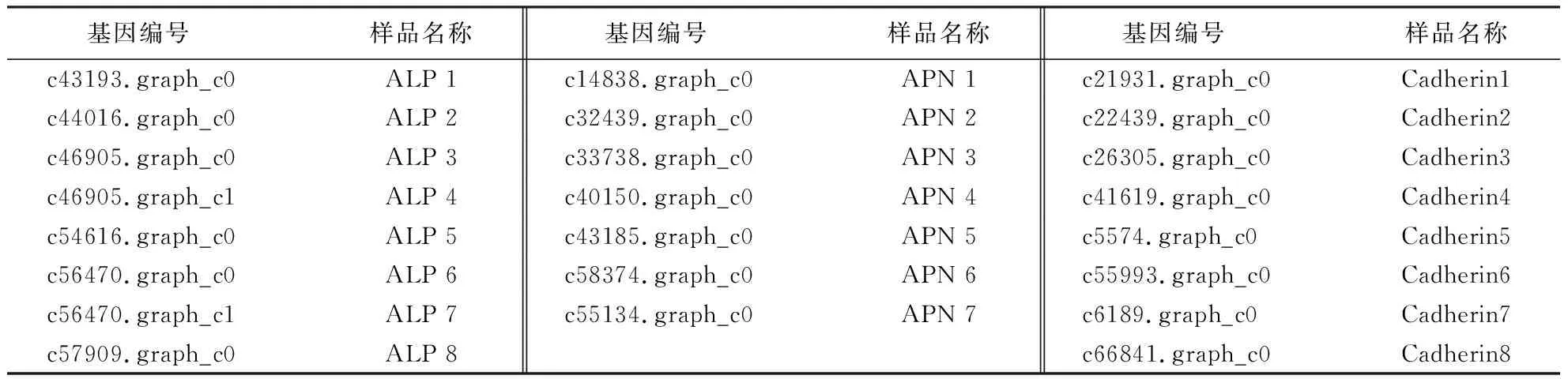
表1 筛选所得的3类受体表达量差异显著的基因及其重命名Table 1 Genes screened with significantly different expression levels in 3 types of receptors and their renamings
2.2 3类受体基因表达量差异的可视化分析
通过Heatmap图可以看到,茶小绿叶蝉肠内外组织大多数ALP、APN、Cadherin基因的相对表达量都有明显的差异,只有少量基因的表达量比较接近.其中,肠外组织中也存在部分表达量相对较高的候选基因,包括ALP 3、ALP 4、ALP 6、ALP 7、ALP 8、APN 7、APN 6、Cadherin6、Cadherin8;在中肠中表达量高的基因有ALP 2、ALP 5、ALP 6、ALP 7、ALP 8、APN 7,其中表达量最高的为APN 7,ALP 8次之(图1).

图中颜色深蓝、淡蓝、黄色、红色(0~8)代表基因表达量逐渐升高的趋势.图1 3类潜在受体基因在肠外组织和中肠中差异表达的Heatmap图Fig.1 Heatmap plots for differential expression of 3 potential receptor genes in intestinal tract and midgut
2.3 RT-PCR分析
将小绿叶蝉RNA反转录为cDNA作模板,以actin基因为内参基因,挑选其中19个在中肠中表达量高且与肠外组织差异表达的ALP、APN、Cadherin基因进行RT-qPCR验证,发现表达相关性系数R2值达到0.951,接近于1(图2).可见,RT-qPCR所计算的表达量数值与RNA-seq基本一致,说明RNA-seq表达量数据准确可靠.

x轴和y轴分别代表RNA-seq和RT-qPCR所计算的基因相对表达量.图2 RNA-seq表达量验证Fig.2 Verification of RNA-seq expression data
2.4 3类潜在受体基因同源性分析
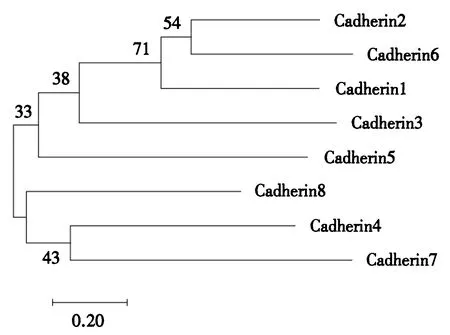
图3 Cadherin受体基因同源性比对进化树Fig.3 Homology alignment of Cadherin receptor genes
通过Mega(Version 7.0)软件构建8个Cadherin潜在受体基因的NJ进化树,发现Cadherin基因可分为2个亚支,Cadherin2、Cadherin6、Cadherin1、Cadherin3、Cadherin5为分支Ⅰ,Cadherin7、Cadherin4、Cadherin8为分支Ⅱ.Cadherin8相对于其他成员较为古老,Cadherin2和Cadherin6可能是一对较新的基因(图3).
利用同种方法构建ALP和APN受体基因的NJ进化树,结果发现,ALP受体基因分别位于3个亚支上,其中ALP 1、ALP 5、ALP 7为分支Ⅰ,ALP 2、ALP 3、ALP 6为分支Ⅱ,ALP 4、ALP 8为分支Ⅲ.ALP 1与ALP 7基因间遗传距离较近.而7个APN基因分别位于2个亚支上,APN 3、APN 5、APN 6、APN 7为分支Ⅰ,APN 1、APN 2、APN 4为分支Ⅱ.
3 讨论
APN、ALP和Cadherin是当前多种昆虫已明确的Cry受体类型,在毒素发挥杀虫活性的过程中起重要作用[14].蚜虫等半翅目昆虫肠道也存在与Bt靶标昆虫类似的ALP、APN等受体,但可能因为其与鳞翅目、双翅目等昆虫的基因同源性较低,导致Cry对其杀虫活性也相对较低[14-15].本研究在前期已获得的茶小绿叶蝉肠内外组织转录组数据的基础上,进一步分析这3类潜在受体的基因信息,通过de novo组装注释的转录本transcripts获得了52 182个组装的Unigene[12],其中有3 327个基因存在差异性表达.同时,从这些差异性表达基因中筛选获得有特异GPI瞄定位点信息的123个显著差异表达的ALP、APN、Cadherin 3类受体基因,包括13个APN、25个ALP、85个Cadherin,其中在中肠中表达量高且中肠内外表达量差异显著的基因有23个,包括7个APN、8个ALP和8个Cadherin.提取茶小绿叶蝉中肠和肠外组织的RNA,通过RT-qPCR分析发现这23个候选基因表达情况与转录组数据分析一致.
为了解茶小绿叶蝉的潜在受体表达量差异情况,选择转录组信息中表达量高且肠内外表达差异显著的23个候选基因为靶标,通过相应的FPKM值得到的相对表达量进行Heatmap作图分析,发现潜在受体基因表达量存在明显差异,其中ALP基因的表达量总体高于APN和Cadherin.而在中肠中表达量高的基因有ALP 2、ALP 5、ALP 6、ALP 7、ALP 8、APN 7,其中APN 7最高,ALP 8次之.综合RNA-seq、RT-PCR及进化树分析可以看出,Cadherin6和Cadherin8表达水平高于其他6个Cadherin类型的受体蛋白,且存在一定的组织特异性,可推测Cadherin6和Cadherin8可能为Bt Cry毒素的潜在受体.在ALP基因中,ALP 1与ALP 7基因间遗传距离较近,但表达量存在差异,ALP 1的表达水平明显低于ALP 7.可见,ALP 1与ALP 7可能存在功能冗余.而在APN基因的表达量比较中发现,APN 7在茶小绿叶蝉中肠中的表达量高于其他潜在受体,且中肠内表达量相对于肠外组织呈指数型增长.因此,APN 7可能为Bt Cry毒素的潜在受体.
综上所述,茶小绿叶蝉存在与鳞翅目、双翅目等Bt靶标昆虫类似的潜在受体,但可能因为自身存在特异性,与其他昆虫同类型受体的同源性相对较低.这些潜在受体与常规受体的差异性是否会影响与毒素结合进而降低Cry毒素的杀虫效果还需进一步深入探讨.
