TSPY基因与肿瘤关系研究进展
2017-12-19刘长海
邵 颖, 刘长海
(延安大学 生命科学学院, 延安 716000)
TSPY基因与肿瘤关系研究进展
邵 颖, 刘长海
(延安大学 生命科学学院, 延安 716000)
睾丸特异性蛋白(testis-specific protein, Y-encoded, TSPY)基因是Y染色体上编码的特异性基因。在人类中,TSPY主要在胎儿和成人睾丸中表达,在胎儿和新生儿发育期间,其表达仅局限于生殖母细胞和前精原细胞中,在成人睾丸中其表达限于精原细胞、初级精母细胞和球形精子细胞。TSPY也在前列腺上皮细胞中微弱表达。TSPY已被归类为在特定器官(睾丸)中高表达的组织富集基因,且在其他组织中不表达或以低水平表达。大量研究发现TSPY基因在人类众多的肿瘤中均特异性表达,揭示出TSPY基因与人类肿瘤的形成过程有着密切的联系。总结阐述TSPY基因在肿瘤形成过程中的功能及其可能的作用机制,旨在对性别依赖性肿瘤的基础研究提供科学参考。
TSPY基因;肿瘤;作用机制
睾丸特异性蛋白(testis-specific protein, Y-encoded, TSPY)基因是一个串联重复基因,每个TSPY基因的拷贝长度约为2.8 kb,具有6个外显子和5个内含子,转录长度约为1.3 kb,编码308个氨基酸[1-3]。TSPY基因于1991年在人类睾丸组织中第一次被发现[4]。Schnieders等[5]通过荧光原位杂交技术将TSPY基因定位在Y染色体短臂近着丝粒处。TSPY基因在人类中拷贝数量为20~76个碱基[6],在健康男性中,TSPY拷贝数可以在个体之间变化,每个基因组的拷贝数在23到64个碱基之间[7],大多数TSPY拷贝位于DYZ5区中一个700 kb的串联重复结构内[8]。有研究表明TSPY基因是一类具有35个基因簇的假基因家族[9],TSPY基因一直被认为是Y染色体上潜在的致癌基因。
1 TSPY基因与肿瘤关系
作为一个潜在的致癌基因,TSPY基因与人类众多肿瘤的形成过程相关,目前发现TSPY基因异变与生殖细胞瘤和各种实体瘤的形成紧密相关,例如,性腺母细胞瘤、前列腺癌、黑色素瘤和肝癌等。
1.1 TSPY基因与生殖细胞瘤
性腺母细胞瘤是一种罕见的生殖细胞肿瘤,主要由原始生殖细胞和性索样细胞组成,其发展和恶化会导致性发育障碍产生[10],如果不加以治疗,性腺母细胞瘤有可能恶化成生殖细胞肿瘤,例如女性的恶性胚胎瘤和男性的精原细胞瘤和非精原细胞瘤。有研究表明,35%的性腺母细胞瘤患者将恶化形成恶性胚胎瘤或精原细胞瘤[11-12]。因此,性腺母细胞瘤被认为是生殖细胞肿瘤的前兆。性腺母细胞瘤的形成与部分基因(TSPY、SPY、WT1、SOX9等)密切相关[13]。在性腺母细胞瘤流行程度的调查研究中,TSPY基因是最常用的候选基因之一[14]。对于TSPY基因表现出阳性的性腺母细胞瘤患者,更有学者推荐采用早期性腺切除术降低其发展为恶性肿瘤的风险[15]。
Zhang等[16]研究发现TSPY基因存在于雄性的基因组中,并且在恒河猴的睾丸中表达,这一发现表明TSPY基因在人类或高等生物中的性腺发育不全等方面具有重要的作用。Tsuchiya等[17]发现在2例不同的性腺母细胞瘤的患者中均有TSPY基因的表达,尽管这一发现并未直接表明TSPY基因是性腺母细胞瘤的发病病因,但却提升了Y染色体上TSPY基因的关键片段或多个片段的表达是性腺母细胞瘤的形成病因的概率。Su等[18]通过超微显微镜技术检测了1例46,XY性腺发育不全患者Y染色体上的14区域内的全部基因,发现只有TSPY基因表达显著,其他基因表达均不显著。他们的发现暗示着TSPY在性腺母细胞瘤的早期形成过程中扮演着重要的角色。Li等[19]将TSPY作为标记研究了4例性腺母细胞瘤的病例发现TSPY基因在样本中都表达,并且TSPY基因在所有的性腺母细胞瘤中都平行表达,同时还发现在性腺母细胞瘤细胞的细胞核和细胞质中均有TSPY存在。他们的研究结果清晰地表明TSPY基因在早期和后期的性腺母细胞瘤和睾丸癌中均表达。Palma等[20]发现TSPY 和OCT3/4是性腺母细胞瘤形成的关键蛋白,他们的研究结果表明TSPY 和OCT3/4的相互作用在性腺母细胞瘤的形成过程中具有重要的作用。同时,TSPY能够作为检测性腺母细胞瘤患病风险的生物学指标。
NG等[21]发现在非精原细胞瘤和大部分的精子发育不良中,TSPY基因表达量下降是TSPY下调的主要原因,而不是Y染色体上相关结构的缺失,这表明TSPY基因表达量降低与非精原细胞瘤的形成具有重要联系。Li等[22]在探索睾丸生殖细胞肿瘤的发病机制中Y染色体上的基因是否参与时,发现TSPY在原位癌、未分化型畸胎瘤和精原细胞瘤中大量表达,而在各类非精原细胞瘤中不表达或以最低限度表达。
1.2 TSPY基因与前列腺癌
前列腺癌是一种常见的恶性肿瘤,也是西方发达国家中男性死亡率第二的癌症。同时,前列腺癌的患病率在亚洲地区也逐年攀升[23]。就目前的研究结果而言,TSPY基因在前列腺癌的形成过程中起着关键作用,在此过程中TSPY基因可能被异常激活,使得前列腺癌的形成表现出异质性[24]。
Lau等[25-26]运用原位mRNA杂交和免疫染色技术证实TSPY优先在前列腺癌病灶区域(高等级和低等级)的上皮细胞中表达,同时还发现雄激素能够上调TSPY基因的表达。这些都表明TSPY可能在经荷尔蒙刺激下的LNCaP细胞分化过程中起作用,这个过程也包含前列腺的癌变过程。Kido等[27-28]曾报道TSPY和真核翻译延伸因子1A(eukaryotic translation elongation factor 1 alpha, eEF1A)共表达时与前列腺癌的形成与恶变过程相关。他们利用免疫组织化学技术研究了41例临床确诊的前列腺癌患者和17例潜在患者样本,发现TSPY的表达与早期肿瘤的形成和癌变相关。而后,在LADY模型动物实验中,利用一种特殊的生物标记物FOXA1(forkhead box protein A1,上皮细胞)证明在肿瘤形成的后期TSPY阳性细胞专一地在肿瘤的基底层增殖和分化。通过对不同分期的前列腺癌临床样本的研究发现,在上皮细胞中,TSPY明显与FOXA1共表达,并且在其他恶性细胞中还与另一种肿瘤标志物AMACR(alpha-methylacyl-CoA racemase)共表达。以上这些研究表明,在人类前列腺癌变的过程中,TSPY基因异常活跃。
1.3 TSPY基因与肝癌
流行病学研究显示,肝癌在全球癌症杀手中位列第3,我国肝癌患病率位居世界第1,居我国癌症死因的第2位。根据2012年出版《五大洲癌症发病率》第Ⅹ卷汇编资料(GLOBOCAN 2012),全球每年肝癌新发病例数为78.25万人(男性55.44万人,女性22.81万人), 位居所有癌症发病的第6位;其中中国每年肝癌新发病例数为39.48万人(男性29.33万人,女性10.15万人)[29]。有研究显示TSPY基因与肝癌的发生具有一定联系。
Yin等[30]利用cDNA 微阵列技术研究与肝癌细胞相关的基因,发现在肝癌细胞中TSPY基因表达量显著上升。Kido等[31]的实验发现TSPY的表达与肝癌的生物标志物(如磷脂酰肌醇蛋白聚糖3)紧密联系。与此相反,TSPX(X染色体上的TSPY同源基因)在肿瘤与非肿瘤的平行样本中(18/33)基因表达量显著下调(-54.5%),这与TSPY和磷脂酰肌醇蛋白聚糖3的表达情况恰恰相反,这也许表明肿瘤形成的过程促使Y染色体上的原癌基因TSPY异常活跃,同时抑制了X染色体上的肿瘤抑制基因TSPX的活性,这也许能解释为什么在肝癌患者中男性偏多的现象。TSPY1(Testis-specific protein Y-encoded 1)特征性的存在于男性肝癌细胞中,其高表达可以促进肝癌细胞的增殖。同时,由于TSPY1和雄激素受体在肝癌细胞群与肝癌细胞组织中的共表达,TSPY1可被用来做男性肝细胞癌的生物标志物[32]。最新的研究结果发现,TSPY基因及其共表达的基因(TSPY co-expression network, TCN)在肝癌中表达量显著上调(30%)[33],揭示了TSPY基因在肝癌的形成过程具有联系。
1.4 TSPY基因与黑色素瘤
黑色素瘤是皮肤癌中最具侵袭性的恶性肿瘤。近20年,世界范围发病率急剧上升,每年以3.1%的速度增加[34]。我国每年新发病例约2万例,近年增长较快[35]。一旦发生转移,预后极差,5年生存率不到10%[36]。通过动物模型研究发现TSPY基因与黑色素瘤的形成具有一定的联系。
Gallagher等[37]系统性地研究了黑色素瘤荷瘤的DAC小鼠,结果发现大量TSPY基因的mRNA表达,这些结果阐明TSPY与黑色素瘤的形成具有一定的联系,同时,TSPY也许是性别特异性标志物,可用于辨别男性和女性之间黑色素瘤发病率的差异性[37]。
1.5 TSPY基因与其他疾病
Kido等研究证实TSPY能够影响神经突触与神经元的传递功能,在体外培养的细胞中,TSPY通过与钙/钙调蛋白依赖的丝氨酸蛋白激酶(calcium/calmodulin-dependent serine protein kinase, CASK)的SET/NAP结合域相结合,从而调节脑部或其他神经组织中的CASK。在神经表达模型动物中,TSPY基因、CASK基因和TSPX基因串联重复,且共同定位于神经轴突纤维。这些研究表明,TSPY与神经系统的发育分化或神经生理具有一定的联系[30]。Kido等[33]通过对肺癌病人的样本研究发现,TSPY基因及其共表达的相关基因在肺癌中表达量显著升高(10%),显示TSPY基因及其共表达的基因有可能参与肺癌的形成过程。
2 TSPY基因作用机制的研究
目前对于TSPY基因的作用机制研究还不是很透彻。相关研究发现TSPY基因功能的呈现与其编码的特殊蛋白质——TSPY有不可分割的关系,并且TSPY能与细胞中的多种物质产生相互作用:1)TSPY能够与组蛋白中的H2A、H2B、H3和H4相结合,在精子形成过程中起重要的作用[38];2)TSPY作为SET/NAP蛋白家族的成员之一,能够与细胞周期蛋白B结合,激活细胞周期蛋白B1-CDK1激酶活性,减少G2期向M期转化的时间,缩短细胞周期,增强细胞更新与分裂的潜能,对细胞的增殖起着重要作用,同样也促进肿瘤细胞的分化与增殖[39];3)TSPY还能与eEF1A相互作用。研究发现,TSPY与eEFlA共同表达于精原细胞瘤中,TSPY可增加eEFlA在细胞核的分布,进而上调细胞的报告基因表达,在这一过程中,TSPY的NAP端的7个氨基酸残基(Asp 175、Glu 176、Asp 179、Tyr 183、Asp 240、Glu 244和Tyr 246)直接与eEF1A上的6个赖氨酸残基结合[40],从而加快与eEFlA有关的细胞进程,促进生殖细胞瘤形成的转录过程,增加蛋白质的合成。同时,在RNA和蛋白水平相同的肿瘤细胞中,TSPY与已确定的生殖细胞肿瘤标志物(PLAP、C-KIT、OCT4)和增殖标记物(Ki-67、cyclin B1)共表达[22]。TSPY基因的异常活跃能够抑制其位于X染色体上的同源基因TSPX基因的活性(TSPX基因被视为抑癌基因)[24],从而导致肿瘤的形成(图1)。
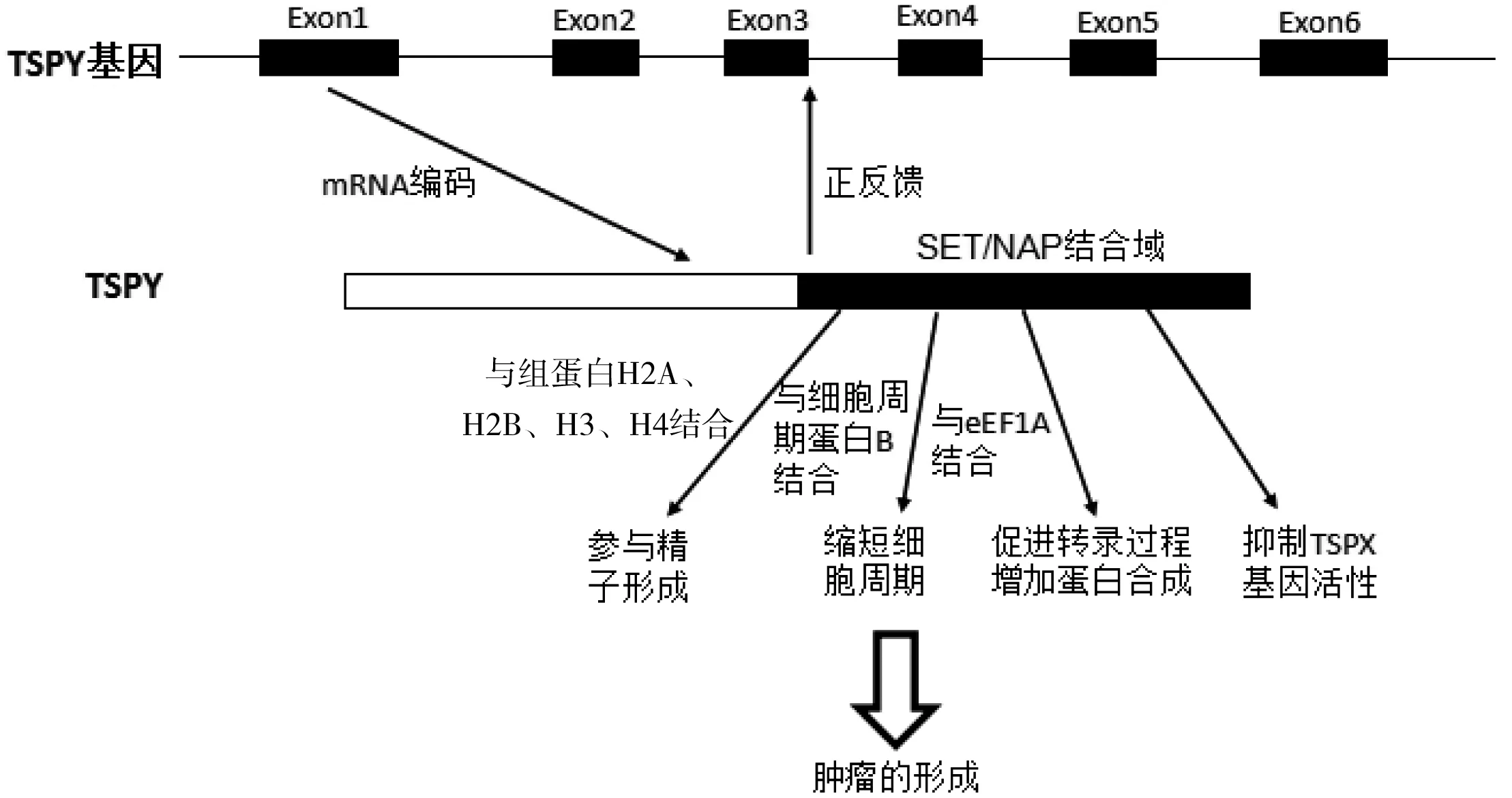
图1 TSPY基因作用机制示意图
TSPY基因在进化中具有高保守性, 人类TSPY基因各拷贝间序列高度同源[40],其中功能拷贝之间的序列差异仅为1%~3%[41-42]。并且,TSPY基因家族能够通过频繁的Y染色体上非等位同源交换导致拷贝数的显著变异。结构基因的拷贝数量的变异与各种重要性状和疾病表型密切相关,结合TSPY基因序列的高保守性以及在生殖细胞瘤和部分实体肿瘤细胞中均特异性表达等性质,可以推断该基因很可能是参与人类精子形成和性别依赖性肿瘤发生的重要功能基因之一。
[1]LAU Y F, LI Y, KIDO T. Gonadoblastoma locus and the TSPY gene on the human Y chromosome [J]. Birth Defects Research(Part C), 2009, 87(1): 114-122.
[2]LAU Y F, LAU H, KÖMÜVES L G. Expression pattern of a gonadoblastoma candidate gene suggests a role of the Y chromosome in prostate cancer [J]. Cytogenetic and Genome Research, 2003, 101(3-4): 250-260.
[3]HONECKER F, STOOP H, DE KRIJGER R R, et al. Pathobiological implications of the expression of markers of testicular carcinoma in situ by fetal germ cells [J]. American Journal of Pathology, 2004, 203(3): 849-857.
[4]AMEMANN J, JAKUBICZKA S, THÜRING S, et al. Cloning and sequence analysis of a human Y-chromosome-derived, testicular cDNA, TSPY [J]. Genomics, 1991, 11(1): 108-114.
[5]SCHNIEDERS F, DÖRK T, AMEMANN J, et al. Testis-specific protein, Y-encoded (TSPY) expression in testicular tissues [J]. Human Molecular Genetics, 1996, 5(11): 1801-1807.
[6]HAMILTON C K, VERDUZCO-GMEZA R, FAVETTA L A, et al. Testis-specific protein Y-encoded copy number is correlated to its expression and the field fertility of Canadian Holstein bulls [J]. Sexual Development, 2012, 6(5): 231-239.
[7]REPPING S, VAN DAALEN S K, BROWN L G, et al. High mutation rates have driven extensive structural polymorphism among human Y chromosome [J]. Nature Genetics, 2006, 38(4): 463-467.
[8]SKALETSKY H, KURODA-KAWAGUCHI T, MINX P J, et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes [J]. Nature, 2003, 423 (6942): 825-837.
[9]SKAWRAN B, SCHUBERT S, DECHEND F, et al. Characterization of a human TSPY promoter [J]. Molecular and Cellular Biochemistry, 2005, 276(1): 159-167.
[10]KATHRINS M, KOLON T F. Malignancy in disorders of sex development [J]. Translational Andrology and Urology, 2016, 5(5): 794-798.
[11]DU X, ZHANG X H, LI Y M, et al. 46, XY female sex reversal syndrome with bilateral gonadoblastoma and dysgerminoma [J]. Experimental and Therapeutic Medicine, 2014, 8(4): 1102-1104.
[12]KAO C S, ULBRIGHT T M, IDREES M T. Gonadoblastoma: an immunohistochem-ical study and comparison to toli cell nodule with intratubular germ cell neoplasia, with pathogenetic implications [J]. Histopathology, 2014, 65(6): 861-867.
[13]YU L L, DONG W R, CHEN M H, et al. Progress in the molecular genetic mechanism of gonadoblastoma [J]. Hereditas, 2015, 37(11): 1105-1115.
[14]MARQUI A B, SILVA-GRECCO R L, BALARIN M A. Prevalence of Y-chromosome sequences and gonadoblastoma in turner syndrome [J]. Revista Paulista de Pediatria, 2016, 34(1): 114-121.
[15]SILVERI M, GROSSI A, BASSANI F, et al. Ullrich-turner syndrome and tumor risk: Is there another chance to early gonadectomy in positive TSPYand SRY Patients? [J]. European Journal of Pediatric Surgery, 2016, 26(3): 271-276.
[16]ZHANG J S, YANG-FENG T L, MULLER U, et al. Molecular isolation and characterization of an expressed gene from the human Y chromosome [J]. Human Molecular Genetics, 1992, 1(9): 717-726.
[17]TSUCHIYA K, REIJO R, PAGE D C, et al. Gonadoblastoma: molecular definition of the susceptibility region on the Y chromosome [J]. American Journal of Human Genetics, 1995, 57(6): 1400-1407.
[18]SU M T, LEE I W , KUO P L. Presence of TSPY transcript and absence of transcripts of other Y chromosomal genes in a case of microscopic gonadoblastoma [J]. Gynecologic Oncology, 2006, 103(1): 357-360.
[19]LI Y, VILAIN E, CONTE F, et al. Testis-specific protein Y-encoded gene is expressed in early and late stages of gonadoblastoma and testicular carcinoma in situ [J]. Urologic Oncology, 2007, 25(2): 141-146.
[20]PALMA I, GARIBAY N, PENA-YOLANDA R, et al. Utility of OCT3/4, TSPY and β-catenin asbiological markers for gonadoblastomaformation and malignant germ cell tumordevelopment in dysgenetic gonads [J]. Disease Markers, 2013, 34(6): 419-424.
[21]NG S B, YONG M H, KNIGHT L A, et al. Gonadoblastoma-associated mixed germ celltumour in 46, XY complete gonadaldysgenesis (Swyer syndrome): analysis of Y chromosomal genotype and OCT3/4 and TSPY expression profile [J]. Histopathology, 2008, 52(5): 644-646.
[22]LI Y M, LAURA T Z, LEE T L, et al. The Y-Encoded TSPY protein: a significant marker potentially plays a role in the pathogenesis of testicular germ cell tumors [J]. Human Pathology, 2007, 38(10): 1470-1481.
[23]ALI-SHAH S I. Emerging potential of parenteral estrogen as androgen deprivation therapy for prostate cancer [J]. South Asian Journal of Cancer, 2015, 4(2): 95-97.
[24]KIDO T, SCHUBERT S, HATAKEYAMA S, et al. Expression of a Y-located human proto-oncogene TSPY in a transgenic mouse model of prostate cancer [J]. Cell and Bioscience, 2014, 4: 9.
[25]LAU Y F. Gonadoblastoma, testicular and prostate cancers, and the TSPY gene [J]. Amreican Journal of Human Genetics, 1999, 64(4): 921-927.
[26]LAU Y F, ZHANG J Q. Expression analysis of thirty one Y chromosome genes in human prostate cancer [J]. Molecular Carcinogenesis, 2000, 27(4): 308-321.
[27]KIDO T, HATAKEYAMA S, OHYAMA C, et al. Expression of the Y-encoded TSPY is associated with progression of prostate cancer [J]. Genes(Basel), 2010, 1(2): 283-293.
[28]KIDO T, LAU Y F. The Y-located gonadoblastoma gene TSPY amplifies its own expression through a positive feedback loop in prostate cancer cells [J]. Biochemical and Biophysical Research Communications, 2014, 446(1): 206-211.
[29]陈建国, 朱 健, 张永辉, 等. 肝癌流行趋势的改变: 基于启东40年癌症登记资料的分析 [J]. 中国肿瘤, 2014, 23(8): 621-628.
[30]YIN Y H, LI Y Y, QIAO H , et al. TSPY is a cancer testis antigen expressed in human hepatocellular carcinoma [J]. British Journal of Cancer, 2005, 93(4): 458-463.
[31]KIDO T, LO R C, LI Y, et al . The potential contributions of a Y-located protooncogene and its X homologue in sexual dimorphisms in hepatocellular carcinoma [J]. Human Pathology, 2014, 45: 1847-1858.
[32]LI S, Mo C J, HUANG S, et al. Over-expressed testis-specific protein Y-encoded 1 as a novel biomarker for male hepatocellular carcinoma [J]. Plos One, 2014, 9(2): e89219.
[33]KIDO T, LAU Y C. Identification of a TSPY co-expression network associated with DNA hypomethylation and tumor gene expression in somatic cancers [J]. Journal of Genetics and Genomics, 2016, 43(10): 577-585.
[34]SPAGNOLO F, QUEIROLO P. Upcoming strategies for the treatment of metastatic melanoma [J]. Archives of Dermatological Research, 2012, 304(3) : 177-184.
[35]斯 璐, 郭 军. 新版中国黑素瘤诊治指南解读 [J]. 临床肿瘤学杂志, 2012, 17(2): 172-173.
[36]LACY K E, KARAGIANNIS S N, NESTLE F O. Advances in the treatment of melanoma [J]. Clinical Medicine, 2012, 12(2): 168-171.
[37]GALLAGHER W M, BERGIN O E, RAFFERTY M, et al. Multiple markers for melanoma progression regulated by DNA methylation: insights from transcriptomic studies [J]. Carcinogenesis, 2005, 26(11): 1856-1867.
[38]KIDO T, LAU Y C. The rat Tspy is preferentially expressed in elongated spermatids and interacts with the core histones [J]. Biochemical and Biophysical Research Communications, 2006, 350(1): 56-67.
[39]STEPHANIE S, JÖRG S. Transgenic mouse studies to understand the regulation, expression and function of the testis-specific protein Y-encoded (TSPY) gene [J]. Genes(Basel), 2010, 1(2): 244-262.
[40]HAMIHON C K, VERDUZCO-GMEZ A R, FAVETTA L A, et al. Testisspecific protein Y-encoded copy number is correlated to its expression and the field fertility of Canadian Holstein bulls [J]. Sexual Development, 2012, 6(5): 231-239.
[41]VOGEL T, SCHMIDTKE J. Structure and function of TSPY, the Y-chromosome gene coding for the "testis-specific protein" [J]. Cytogenetics and Cell Genetics, 1998, 80(1-4): 209-213.
[42]SKAWRAN B, SCHUBERT S, DECHEND F, et al. Characterization of a human TSPY promoter [J]. Molecular and Cellular Biochemistry, 2005, 276(1): 159-167.
ProgressoftherelationshipbetweenTSPYgeneandhumantumor
SHAO Ying, LIU Chang-hai
(College of Life Science, Yan′an University, Yan′an 716000, China)
Testis-specific protein, Y-encoded (TSPY) gene is a specific gene which is located and encoded on Y chromosome. In humans, TSPY is mainly expressed in fetal and adult testis, where its expression is limited to gonocytes and prespermatogonia during fetal and neonatal development, and restricted to spermatogonia, primary spermatocytes and round spermatids in adult testis. TSPY is also expressed, although much more weakly, in epithelial cells of the prostate gland. TSPY has been classified as a tissue-enriched gene that is highly expressed in a specific organ (testis) and is not expressed, or is expressed at much lower levels, in other tissues. It was found in a large number researches that specific expression of TSPY gene exists in many human tumors, which revealed a fact that TSPY gene is closely related to the formation of human tumors. The function and possible mechanism of TSPY gene in the process of tumor formation will be discussed in order to provide scientific reference for the basic research of sex-dependent tumors.
TSPY gene; human tumor; the mechanism of TSPY gene
2016-11-14;
2016-12-20
国家自然科学基金项目(31370541);陕西省自然科学基金项目(2013JM3018);陕西省高水平大学建设专项资金资助项目(2012SXTS03)
邵 颖,硕士研究生,主要从事动物生态学研究,E-mail:1076958605@qq.com
刘长海,博士,教授,主要从事动物生态学教学与研究,E-mail:yadxlch@126.com
10.3969/j.issn.2095-1736.2017.06.069
R730.2
A
2095-1736(2017)06-0069-04
