急性T淋巴细胞白血病相关长链非编码RNA T-ALL-R-LncR1的发现、鉴定及其功能研究
2017-12-12张琳陆超李天宇
张琳,陆超,李天宇
(1无锡市儿童医院,江苏无锡214023;2江苏省人民医院)
急性T淋巴细胞白血病相关长链非编码RNA T-ALL-R-LncR1的发现、鉴定及其功能研究
张琳1,陆超2,李天宇1
(1无锡市儿童医院,江苏无锡214023;2江苏省人民医院)
目的对新发现的急性T淋巴细胞白血病(T-ALL)相关长链非编码RNA进行鉴定并研究其功能。方法①提取T-ALL Jurkat细胞RNA进行全转录组深度测序,发现了一种新的T-ALL相关长链非编码RNA,命名为T-ALL-R-LncR1。用实时荧光定量PCR和琼脂糖凝胶电泳法观察Jurkat细胞和T-ALL患者骨髓单个核细胞中T-ALL-R-LncR1的表达情况。②将体外培养的T-ALL患者骨髓单个核细胞分为实验组、空质粒组和对照组。实验组转染T-ALL-R-LncR1小干扰RNA(siRNA),空质粒组转染空质粒,对照组不转染。转染24 h时用实时荧光定量PCR法检测各组细胞T-ALL-R-LncR1相对表达量。转染24、48、72 h时用CCK-8法观察各组细胞增殖情况(以OD450表示),转染48 h用流式细胞术测算各组细胞凋亡率。结果① T-ALL-R-LncR1具有原癌基因特性,在人T-ALL Jurkat细胞及T-ALL患者骨髓单个核细胞中高表达。用逆转录实时定量PCR法从Jurkat细胞及急性T淋巴细胞白血病骨髓单个核细胞中获得PCR产物的琼脂糖凝胶电泳结果符合T-ALL-R-LncR1。测序结果证实PCR产物为T-ALL-R-LncR1。②实验组、空质粒组和对照组T-ALL-R-LncR1相对表达量分别为0.79±0.18、 1.12±0.08和1。实验组骨髓单个核细胞 T-ALL-R-LncR1相对表达量低于空质粒组和对照组(P均<0.05),空质粒组和对照组骨髓单个核细胞 T-ALL-R-LncR1相对表达量相比P>0.05。转染24、48、72 h时实验组骨髓单个核细胞OD450值均低于空质粒组和对照组(P均<0.05),空质粒组和对照组骨髓单个核细胞OD450相比P均>0.05。转染48 h时实验组、空质粒组、对照组细胞凋亡率分别为38.67%±1.32%、1.80%±0.14%、1.79%±0.13%。实验组细胞凋亡率高于空质粒组和对照组(P均<0.05),空质粒组和对照组细胞凋亡率相比P>0.05。结论本研究发现了一种具有原癌基因特性的新T-ALL相关长链非编码RNA,即T-ALL-R-LncR1。沉默T-ALL-R-LncR1可以促进T-ALL患者骨髓单个核细胞凋亡并抑制其增殖。
急性T淋巴细胞白血病;长链非编码RNA;细胞增殖;细胞凋亡
急性T淋巴细胞白血病(T-ALL)在儿童血液系统恶性肿瘤中的发病率约为15%,病死率约20%[1]。目前T-ALL的发病机制仍不清楚。研究发现,长链非编码RNA(LncRNA)参与了肿瘤浸润与转移、细胞凋亡调控等过程,并可通过表观遗传调控的方式影响肿瘤细胞生长,可能成为新型肿瘤标志物或肿瘤治疗的靶点[2,3]。为寻找T-ALL相关LncRNA,本研究于2012年3月对T-ALL细胞株Jurkat进行全转录组深度测序,将新发现的T-ALL相关LncRNA命名为T-ALL-R-LncR1,并观察了转染T-ALL-R-LncR1 siRNA对T-ALL患儿骨髓单个核细胞增殖、凋亡的影响,旨在明确T-ALL-R-LncR1在T-ALL中的作用及其机制。
1 材料与方法
1.1 Jurkat细胞株来源及T-ALL患儿骨髓单个核细胞制备 Jurkat细胞系购于中国科学院上海生命科学研究院细胞资源中心。抽取无锡市儿童医院血液科收治的1例T-ALL患儿的骨髓2 mL,肝素抗凝,用淋巴细胞Ficoll分离液分离骨髓单个核细胞,用含100 g/L FBS的RPMI 1640培养基于37 ℃、50 mL/L CO2细胞培养箱中培养。细胞呈悬浮状态,生长良好,处于对数生长期时进行后续实验。本研究已获我院医学伦理委员会批准,患儿家长签署知情同意书。
1.2 T-ALL相关LncR的检测及鉴定 收集处于对数生长期的Jurkat细胞,接种于细胞培养板,每孔106个细胞。每孔加入1 mL TRIzol裂解细胞,用移液器(去RNA酶)轻轻来回吹吸裂解细胞,将裂解后的细胞全部转移至无菌无RNA酶的1.5 mL EP管,置于干冰箱中运送至深圳华大基因公司进行全转录组深度测序。测序数据由华大基因公司进行生物信息分析,获得一种T-ALL相关的长链非编码RNA,并命名为T-ALL-R-LncR1。采用实时荧光定量PCR法检测T-ALL-R-LncR1在Jurkat细胞及FALL患者骨髓单个核细胞中的表达情况。引物设计应用Primer5.0软件。T-ALL-R-LncR1上游引物序列:5′-TGCCAGCACGGCTAGGATAAA-3′,下游引物序列:5′-GTGAAGGAAGCCTGGTTTTGA-3′;内参照GAPDH上游引物:5′-GGTCGGAGTCAACGGATT-TGGTCG-3′,下游引物5′-CCTCCGACGCCTGCTTCA-CCAC-3′。PCR扩增反应条件:98 ℃预变性1 min,然后98 ℃变性30 s、64 ℃退火30 s、72 ℃延伸2 min,热循环(变性、退火和延伸)35次,最后72 ℃延伸7 min,4 ℃保存。12 g/L琼脂糖凝胶电泳检测PCR产物大小,部分PCR产物送上海英骏生物技术有限公司测序。
1.3 转染T-ALL-R-LncR1 siRNA对T-ALL患儿骨髓单个核细胞增殖、凋亡的影响 将体外培养的T-ALL患儿骨髓单个核细胞接种于6孔培养板,每孔1.5 mL培养基含(3~5)×105个细胞。设实验组、空质粒组和对照组,每组6孔。实验组转染T-ALL-R-LncR1小干扰RNA(siRNA),空质粒组转染空质粒,对照组不转染。转染方法参照LipofectamineTM2000说明书。
1.3.1 各组骨髓单个核细胞T-ALL-R-LncR1相对表达量检测 采用实时荧光定量PCR法。转染24 h后收集各组细胞,采用TRIzol一步法提取各组细胞总RNA,反转录为cDNA。行实时荧光定量PCR检测。T-ALL-R-LncR1上游引物序列5′-GAAGGCGCCATTTCTCTGTG-3′,下游引物序列5′-AGATGTGCGGGTCTTTTCCG-3′;内参照β-actin上游引物序列5′-ACTGGAACGGTGAAGGTGAC-3′,下游引物序列5′-AGAGAAGTGGGGTGGCTTTT-3′。反应体系为10 μL。PCR扩增条件:95 ℃预变性30 s,95 ℃变性5 s、60 ℃退火30 s,共40个循环,最后95 ℃ 15 s、60 ℃ 1 min、95 ℃ 15 s。采用2-ΔΔCt法计算T-ALL-R-LncR1相对表达量。
1.3.2 各组骨髓单个核细胞增殖情况观察 采用CCK-8法。转染24、48、72 h时每孔加入10 μL CCK-8,轻轻混匀,37 ℃条件下孵育2 h,用全自动酶标仪检测450 nm处的吸光度(OD450)值,用其表示各组骨髓单个核细胞增殖情况。
1.3.3 各组骨髓单个核细胞凋亡率测算 采用流式细胞术。转染48 h,收集各组细胞悬液至2.0 mL的EP管,1 000 r/min离心5 min,弃上清。用预冷的PBS缓冲液洗涤细胞2次,调整细胞密度至1×105/mL,100 μL细胞悬液中加5 μL AnnexinV-APC和5 μL放线菌素D(7AAD),混匀,避光,室温放置15 min后每个样品加入400 μL Binding buffer,振荡混匀后用流式细胞仪检测凋亡率。
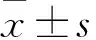
2 结果
2.1 T-ALL-R-LncR1的特征及其在Jurkat细胞、T-ALL骨髓单个核细胞中的表达情况 T-ALL-R-LncR1由2个外显子组成,2个外显子的大小分别是338、1 130 bp,这2个外显子在6号染色体上的起点和终点分别是147176058、147177525,使用RNAFold软件对T-ALL-R-LncR1进行了深层次的分析,发现其有稳定的二级结构,但是在其开放阅读框架(ORFs)5′端的转录起始位点缺乏Kozak序列,它的ORFs不能翻译成蛋白质。用逆转录实时定量PCR法从Jurkat细胞及T-ALL骨髓单个核细胞中获得PCR产物琼脂糖凝胶电泳结果符合T-ALL-R-LncR1。送上海英骏生物技术有限公司测序的PCR产物证实为T-ALL-R-LncR1。
2.2 各组骨髓单个核细胞T-ALL-R-LncR1相对表达量比较 实验组、空质粒组和对照组T-ALL-R-LncR1相对表达量分别为0.79±0.18、 1.12±0.08和1。实验组骨髓单个核细胞 T-ALL-R-LncR1相对表达量低于空质粒组和对照组(P均<0.05),空质粒组和对照组骨髓单个核细胞 T-ALL-R-LncR1相对表达量相比P>0.05。
2.3 各组骨髓单个核细胞增殖情况比较 相同时间点实验组骨髓单个核细胞OD450值低于空质粒组和对照组(P均<0.05),空质粒组和对照组骨髓单个核细胞OD450值相比P>0.05。见表1。

表1 各组转染24、48、72 h时骨髓单个核细胞OD450值比较
注:与对照组、空质粒组相比,*P均<0.05。
2.4 各组骨髓单个核细胞凋亡率比较 转染48 h时实验组、空质粒组、对照组细胞凋亡率分别为38.67%±1.32%、1.80%±0.14%、1.79%±0.13%。实验组细胞凋亡率高于空质粒组和对照组(P均<0.05),空质粒组和对照组细胞凋亡率相比P>0.05。
3 讨论
LncRNA是一类转录本长度200 nt~100 kb的非编码RNA,其表达具有时空特异性,是不具备编码蛋白功能的基因转录产物[4]。目前越来越多的研究证实,LncRNA以RNA的形式在表观遗传调控、转录调控及转录后调控等多种层面调控基因的表达水平,它既可抑制蛋白编码基因的表达,也可活化某些蛋白编码基因表达。其作用机制亦多种多样,如介导染色质重构及组蛋白修饰、影响下游基因的表达、影响转录因子活化与转运、调节特定蛋白质的活性等[5]。多种肿瘤中存在LncRNA的异常表达,尤其是恶性肿瘤[6]。在肿瘤细胞中,某些特定LncRNA的表达水平会发生特异性改变,在结肠癌细胞中OCC1基因的转录水平明显增高, OCC1 RNA具有明显的组织特异性,且在正常黏膜中无或很少表达[7]。在前列腺癌中,DD3RNA只在前列腺组织中被特异性转录,约90%的前列腺癌患者中存在其转录产物水平增高[8],故DD3RNA被作为特异性的标志物用于前列腺癌的诊断[9]。在小细胞肺癌组织中MALAT-1与肺癌转移形式的特殊关联使其成为肺癌患者诊断的标志物[10]。HOTAIR与乳腺癌、肝癌的复发、转移有关[11]。Eis等[12]研究发现,miR-155可由淋巴细胞中LncRNA BIC RNA序列加工而成。但目前为止,临床上对T-ALL相关LncRNA的研究较少。
本研究我们通过高通量测序获得了一个新的T-ALL相关LncRNA,将其命名为T-ALL-R-LncR1。T-ALL-R-LncR1由2个外显子组成,2个外显子的大小分别是338、1 130 bp,这2个外显子在6号染色体上的起点和终点分别是147176058、147177525,使用RNAFold软件对T-ALL-R-LncR1进行深层次的分析发现,其有稳定的二级结构,但是在其开放阅读框架(ORFs)5′端的转录起始位点缺乏Kozak序列,它的ORFs不能翻译成蛋白质。这些表明T-ALL-R-LncR 具有原癌基因特点。本研究用实时荧光定量PCR和琼脂糖凝胶电泳法观察Jurkat细胞和T-ALL患者骨髓单个核细胞中T-ALL-R-LncR1的表达情况,结果电泳结果符合T-ALL-R-LncR1。送去测序的PCR产物也被证实为T-ALL-R-LncR1。表明T-ALL患儿骨髓单个核细胞中存在T-ALL-R-LncR1表达,T-ALL-R-LncR1可能是T-ALL的相关LncRNA。
为进一步明确T-ALL-R-LncR1与T-ALL的相关性,并研究T-ALL-R-LncR1在T-ALL发生发展中的作用,本研究观察了转染T-ALL-R-LncR1 siRNA对T-ALL患儿骨髓单个核细胞增殖、凋亡的影响,结果显示,相对于转染空质粒的空质粒组和未转染的对照组,转染T-ALL-R-LncR1 siRNA的实验组T-ALL患儿骨髓单个核细胞T-ALL-R-LncR1相对表达量降低,细胞凋亡率升高,细胞增殖能力下降。表明T-ALL-R-LncR1确实参与了T-ALL患儿骨髓单个核细胞增殖和凋亡的调控。
搜索国内外数据库,笔者并未发现类似内容的文献。故本研究发现了一个新的与T-ALL有关的LncRNA:T-ALL-R-LncR1;沉默T-ALL-R-LncR1能抑制T-ALL患儿骨髓单个核细胞增殖,促进T-ALL患儿骨髓单个核细胞凋亡。T-ALL-R-LncR1是T-ALL治疗的一个潜在的靶点。但是本研究并未涉及T-ALL-R-LncR1调控T-ALL患儿骨髓单个核增殖、凋亡的机制,今后我们将进行这方面的研究。
[1] Van Vlierberghe P, Ferrando A. The molecular basis of T cell acute lymphoblastic leukemia[J]. Clin Invest, 2012,122(10):3398-3406.
[2] Brockdorff N. Noncoding RNA and polycomb recruitment[J]. RAN, 2013,19(4):429-442.
[3] Zhang L, Xu HG, Lu C. A novel long non-coding RNAT-ALL-R-LncR1 knockdown and Par-4 cooperate to induce cellular apoptosis in T-cell acute lymphoblastic leukemia cells[J]. Leukemia Lymphoma, 2014,55(6):1373-1382.
[4] Mercer TR, Dinger ME, Mattick JS, et al. Long non-coding RNAs: insights into functions[J]. Nat Rev Genet, 2009,10(3):155-159.
[5] Wilusz JE, Sunwoo H,Spector DL. Long noncoding RNAs: functional surprises from the RNA world[J]. Genes Dev, 2009,23(13):1494-1504.
[6] Fu X, Ravindranath L, Tran N, et al. Regulation of apoptosis by a prostate-specific and prostate cancer-associated nonconding gene PCGEM1[J]. DNA Cell Biol, 2006,25(3):135-141.
[7] Pibouin L, Villaud YJ, Ferbus D, et al. Cloning of the mRNA of overexpression in colon carcinoma-1: a sequence overexpressed in a subset of colon carcinomas[J]. Cancer Gen Cytog, 2002,133(1):55-60.
[8] Bussemakers MJC, Van Bokhoven A, Verhaegh GW, et al. DD3: a new prostate-specific gene, highly overexpressed in prostate cancer[J]. Cancer Res, 1999,59(23):5975-5979.
[9] De Kok JB, Verhaegh GW, Roelofs RW, et al. DD3PCA3, a very sensitive and specific marker to detec prostate tumors[J]. Cancer Res, 2002,62(9):2695-2698.
[10] Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel nonconding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer[J]. Oncogene, 2003,22(2):8031-8041.
[11] Niinuma T, Suzuki H, Nojima M, et al. Upgulation of miR-196a and HOTAIR drive malignant character in gastrointestinal stromal tumors[J]. Cancer Res, 2012,72(5):1126-1136.
[12] Eis PS, Tam W, Sun L, et al. Accumulation of miR-155 and BIC RNA in human B cell lymphomas[J]. Proc Natl Acad Sci U S A, 2005,102(10):3627-3632.
国家自然科学基金资助项目(81170487);无锡市卫生计生委“科教强卫工程”医学青年人才项目资助(QNRC014);无锡市卫生局重大项目(Z201302)。
李天宇(E-mail: wxlty999@163.com)
10.3969/j.issn.1002-266X.2017.40.022
R733.71
B
1002-266X(2017)40-0042-03
2016-08-11)
