川东南小型水库营养结构特征的稳定C、N同位素分析
2017-11-29王志坚徐丹丹王永明
李 斌 王志坚 徐丹丹 王永明 陶 敏
(1. 内江师范学院生命科学学院, 内江 641100; 2. 淡水鱼类资源与生殖发育教育部重点实验室, 西南大学, 重庆 400715)
川东南小型水库营养结构特征的稳定C、N同位素分析
李 斌1,2王志坚2徐丹丹1王永明1陶 敏1
(1. 内江师范学院生命科学学院, 内江 641100; 2. 淡水鱼类资源与生殖发育教育部重点实验室, 西南大学, 重庆 400715)
为了解我国水库生态系统营养结构特征, 研究应用稳定同位素技术分析了四川省东南部的典型小型水库松林水库中不同水生生物碳、氮稳定性同位素比值。基于Bayesian混合模型(SIAR)分析了不同消费者基础碳来源, 并计算了δ13C–δ15N同位素生态位中6个营养结构量化指标。结果表明: 调查期间松林水库处于富营养化状态; 初级生产者POM(主要成分为浮游藻类)、固着藻类、喜旱莲子草Alternanthera philoxeroides和水蓼Polygonum hydropiper的δ13C值范围为–29.20‰—18.81‰,δ15N值范围为4.01‰—12.73‰; 其中POM和固着藻类是多数消费者的主要碳源; 松林水库食物网营养级长度为3级, 以杂食性鱼类为优势类群并存在营养冗余现象, 暗示了该生态系统鱼类群落结构的相对稳定性; 入侵物种福寿螺Pomacea lineata、罗非鱼Oreochromisspp.与土著物种铜锈环棱螺Bellamya aeruginosa、鲫Carassius auratus等之间存在明显的同位素生态位重叠现象。建议加强水生生物资源管理, 减少外来物种入侵对当地土著物种的保护具有重要意义。
水库; 营养结构; 碳、氮同位素; 同位素生态位; Bayesian混合模型
水库在经济社会发展中起重要作用:提供生活及工农业水源,防洪发电,区域生态服务,推动渔业产业发展等[1,2]。在日益频繁的人类活动的影响下,库区及周边地区污染物的输入往往导致水体富营养化、生物多样性下降等一系列生态系统退化现象的产生[3]。水库生态系统营养结构遭到破坏,其服务功能也随之下降或丧失[4]。因此,开展水库生态系统营养关系的研究是有效保护和管理水库的重要前提。
近年来,稳定碳、氮同位素技术被广泛用于探究不同水域生态系统营养结构特征并获得较好的研究结果[5]。该方法利用营养物质在食物链传递过程中稳定碳、氮同位素的分馏规律(即消费者δ13C和δ15N值与其食物相比富集度分别约为0—1‰和3.4‰)来确定食物来源[5,6]及消费者营养级水平[7,8]。目前,稳定性同位素技术在世界范围内已经得到广泛应用,在我国多被用于湖泊、河流和海洋生态系统研究[9—11],而水库生态系统罕见相关报道。
松林水库位于四川省东南部内江市东兴区,是当地重要的饮用水源,也是川东南众多小型水库的典型代表。本研究以松林水库为例,应用稳定碳、氮同位素技术对该水域生态系统基础碳来源、能量流通途径、食物链长度及物种之间的营养关系进行探究,以期为松林水库环境管理和渔业利用提供科学依据,也为我国水库生态学研究积累新的资料。
1 材料与方法
1.1 研究区域概况
松林水库位于我国四川省东南部内江市东兴区。该地区属于亚热带季风气候,年平均气温15—28℃;降雨量约1000 mm,主要集中在6—9月;地势为浅丘平坝相间,是典型的川中丘陵地貌。该水库是一座以灌溉为主,兼防洪、供水和水产养殖等功能为一体,总库容1453万m3,年供水量超过50万m3[12],周边有大量的耕地、农田和民居。本研究采样点设置(图1)。
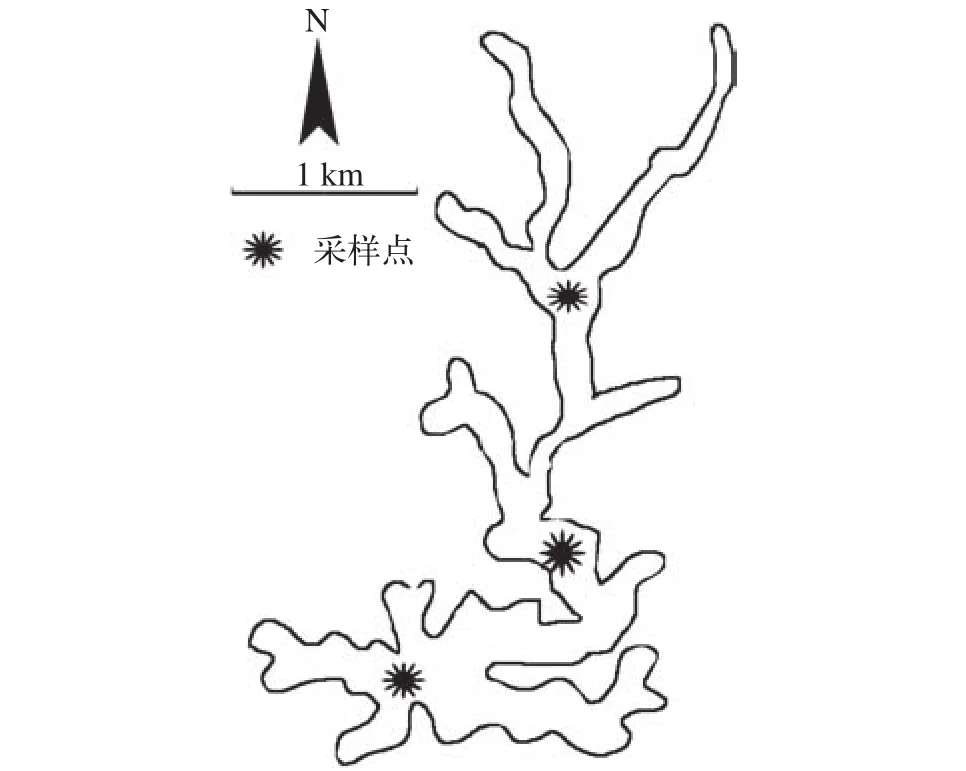
图1 采样点分布图Fig. 1 Location of the Songlin Reservoir and sampling sites
1.2 样品的采集与处理
理化指标测定于2016年8月使用5 L辛德勒采水器采集松林水库表层水样,密封冷藏运回实验室。水体透明度(Sec-chi Depth, SD)用塞氏盘现场度量。使用便携式水质分析仪器(YSI-550A等)原位测定水温、pH和溶解氧(DO)。其余水化学指标,包括总氮(TN)、氨氮(NH4-N)、硝态氮(NO3-N)、正磷酸盐(PO4-P)以及化学需氧量(CODMn)的测定参照文献[13]。叶绿素a测定参照金相灿等[14]和章宗涉等[15]方法,其计算公式为:Chl.a丙酮=27.9×[(E665–E750)–(A665–A750)]×V丙酮/V水样。
同位素样品收集与处理于2016年8月徒手采集松林水库沿岸优势水生植物,用清水除去表面附着物。水中颗粒有机物(POM,经过鉴定主要成分为浮游藻类)采集方法如下:取水库表层水,先经浮游动物网过滤,将滤液再用玻璃纤维滤膜(GF/C Whatman,已预烧450℃)过滤,最后把含有颗粒有机物样品的滤膜用锡箔纸包裹,存放于封口袋中(每一个POM样品至少由3个相应的样混合而成)。利用三角形手抄网(100目)采集石蚕Phryganea japonica、箭蜓Gomphidae等底栖动物;徒手采集福寿螺Pomacea lineata和铜锈环棱螺Bellamya aeruginosa,分别放置于充分曝气的蒸馏水中过夜,使其肠含物排空,然后去除外壳获取软组织。每个采样点采集的底栖动物样品均由多个个体混合而成,样品数量须能满足同位素测试需要。用电鱼机采集鱼类样品,取其背侧肌肉存放在准备好的离心管中。所有样品均密封冷藏运回实验室进行预处理,不能及时处理的样本于–20℃冷冻保存备用。用于碳稳定性同位素分析的滤膜先用1 mol/L的盐酸处理,然后用蒸馏水冲洗干净。最后将所有样品在60℃烘干至恒重,用研钵研磨成粉末,放入干燥器保存待测。
1.3 同位素测试
实验样品的稳定C、N同位素比值在北京林业科学院同位素实验室用稳定同位素质谱仪测定,该仪器由元素分析仪(菲尼根Flash EA112 HT)与同位素比率质谱仪(菲尼根DELTA V Advantage)相连而成。
稳定C、N同位素的自然丰度表示为:δX=(R样品/R标准–1)×1000‰
式中:X为13C或15N ,R为13C/12C或15N/14N。δ13C值是相对于国际PDB标准,δ15N值是相对于空气氮气的丰度。分析精度δ13C<0.10‰,δ15N<0.20‰。
1.4 数据统计与分析
营养级水平计算公式[16]:TP (Trophic position)=[(δ15Nconsumer–δ15Nbaseline)/Δδ15N‰]+λ
式中,TP为营养级水平;δ15Nconsumer为消费者氮稳定同位素值;δ15Nbaseline为基准物种氮稳定同位素值;Δδ15N=3.4‰;当baseline是生产者时,λ=1;当baseline初级消费者是,λ=2。
Bayesian混合模型分析[17]参考McCutchan等[18]和Mao等[10],应用R软件(Ri386 3.3.0)分析松林水库中4种初级生产者POM、固着藻类、喜旱莲子草和水蓼对不同消费者碳源的贡献比例。
采用6个指标:氮值范围NR (δ15N rang)、碳值范围CR (δ13C rang)、总面积TA (Total area)、平均离心距离CD (Mean distance to centroid)、平均最邻近距离NND (Mean nearest neighbor distance)、最邻近距离标准差SDNND (Standard deviation of nearest neighbor distance)用于量化营养结构[19]。其中TA计算利用ArcGIS 9.3软件完成,其余指标计算均利用Excel 2003软件完成。统计分析部分借助STATISTICA.6.0和Spss16.0软件实现。
2 结果
2.1 理化指标
从表1可知,松林水库溶解氧变化范围为8.50—9.60 mg/L,pH为7.60—7.80 mg/L,化学需氧量(CODMn)为11.78—12.36 mg/L;磷酸盐()均值为(0.13±0.15) mg/L,总氮(TN)为(3.519±0.033) mg/L,超出饮用水源地水质标准,为劣于V类状态(GB3838-2002)。
2.2 初级生产者同位素组成
松林水库生主要初级生产者为POM(经鉴定主要为浮游藻类)、底栖固着藻类、沿岸优势水生植物喜旱莲子草A. philoxeroides与水蓼P. hydropiper(表2)。其中,以固着藻类δ13C值最大,为–19.09‰±0.28‰;喜旱莲子草和水蓼的δ13C值差异不大,分别为–28.13‰±0.45‰和–28.89‰±0.31‰。δ15N值组成方面,以固着藻类δ15N值最高(12.73‰)(表2);POM样品δ15N值最低(平均值为4.15‰±0.14‰)。浮游藻类和固着藻类的C∶N值分别为7.66±0.98和3.54±0.7。
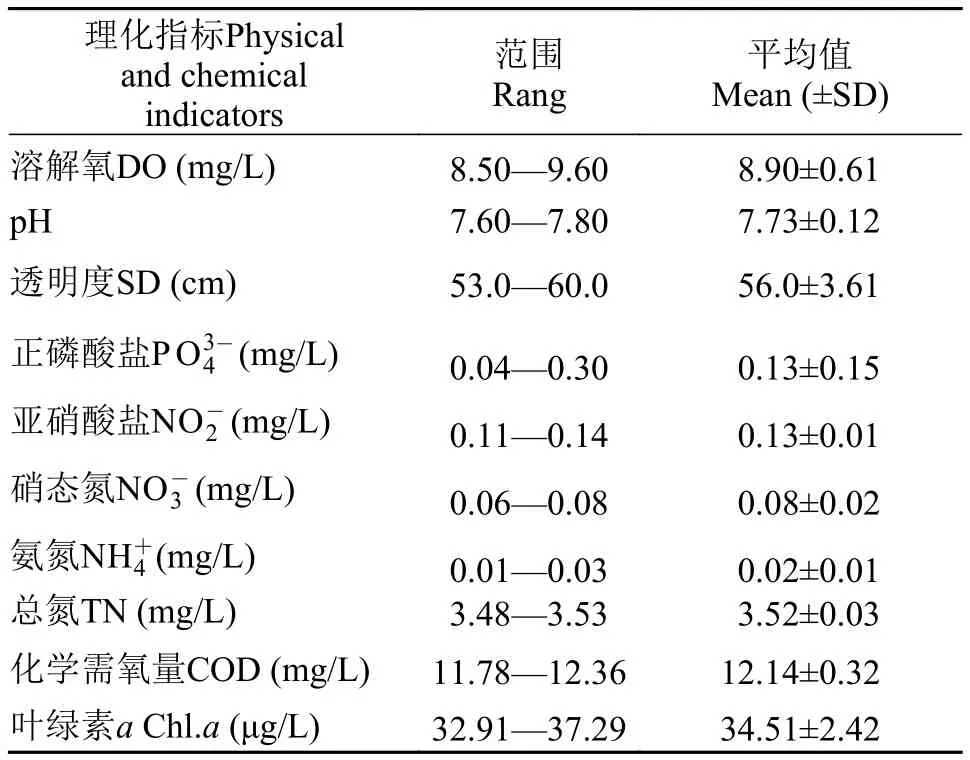
表1 松林水库理化指标Tab. 1 Mean (±SD) summary of physical and chemical variables in the Songlin Reservoir
2.3 消费者同位素组成
在消费者的δ13C值方面,底栖无脊椎动物石蚕δ13C值最低,为–28.90‰,食蚊鱼Gambusiaspp.δ13C值最大,范围为–21.91‰— –20.90‰。δ15N值方面,以黑尾Hemiculter tchangi δ15N值最大为12.28‰±1.28‰ (平均值);无脊椎动物铜锈环棱螺δ15N值最低为6.09‰±1.45‰ (平均值)。方差分析(ANOVA)显示,不同底栖动物物种δ13C值之间(P=0.418,F=1.15,SS=14.77,MS=2.96)差异不明显,但其δ15N (P<0.01,F=46.1,SS=32.27,MS=4.61)值组成差异显著;不同鱼类δ13C之间(P=0.549,F=0.94,SS=35.73,MS=4.47)、δ15N(P=0.076,F=2.95,SS=39.98,MS=7.99)差异均不显著。本研究中所有消费者的C∶N比值介于2.04—5.72。
2.4 营养级水平
通常在水域生态系统中不同营养级间的δ15N值平均富集度为3.4‰[16],依据Post[16]的建议,本研究中选择铜锈环棱螺(土著物种)的δ15N值作为基线值计算了其他消费者的营养级水平(表2)。如表2、图2所示, 松林水库营养级长度约为3(3.8)级,底栖无脊椎动物除秀丽白虾Palaemon modestus位于第三营养级外,其他都位于第二营养级,分布也相对松散;不同鱼类的营养级介于3.09—3.82,其中以黑尾最高(3.82),位于食物网顶端(图2)。
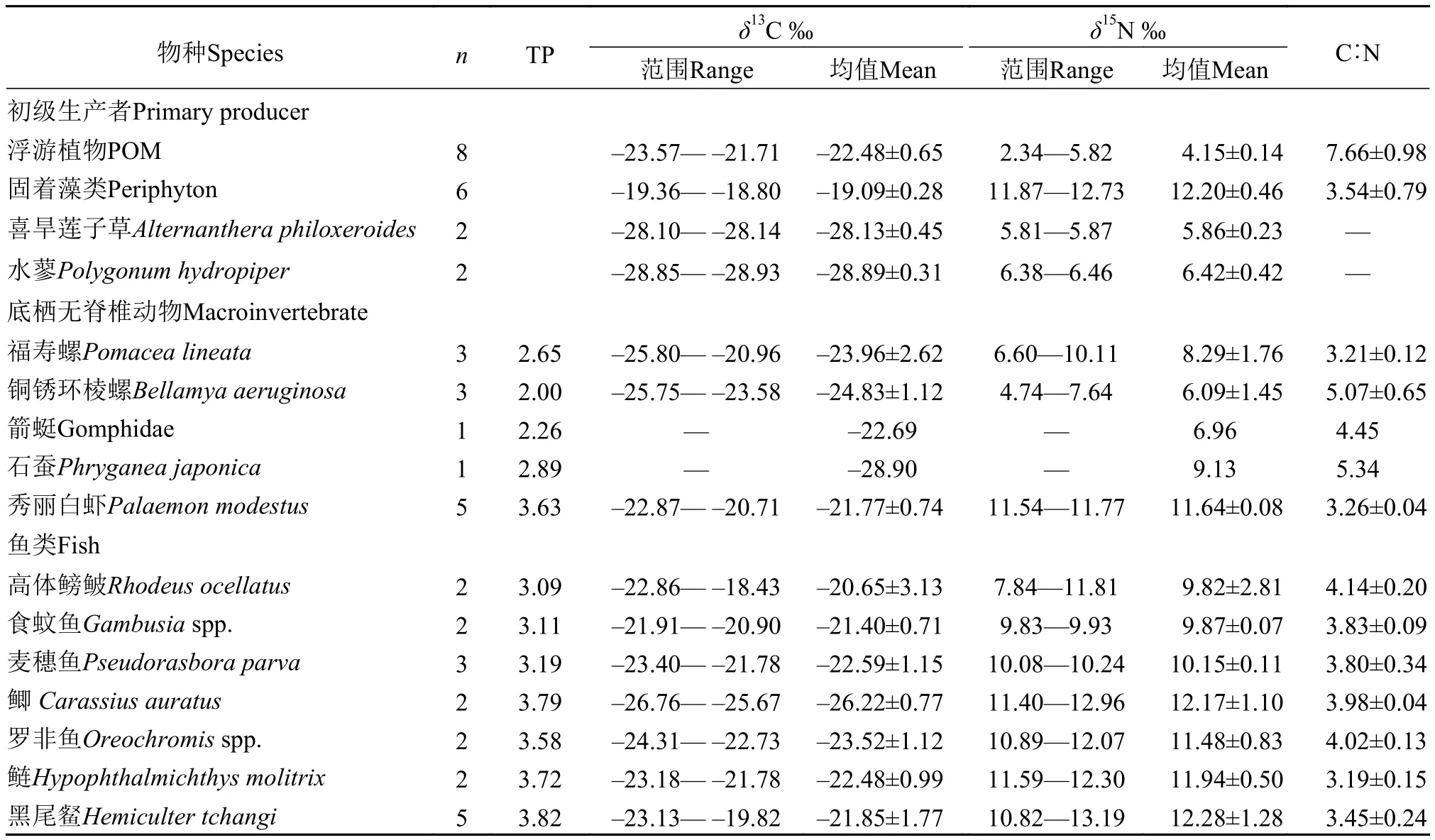
表2 松林水库不同水生生物碳、氮同位素值组成(n为样本量)Tab. 2 Rang and mean (±SD) of the δ13C and δ15N ratios of the Songlin Reservoir (n=sample size)
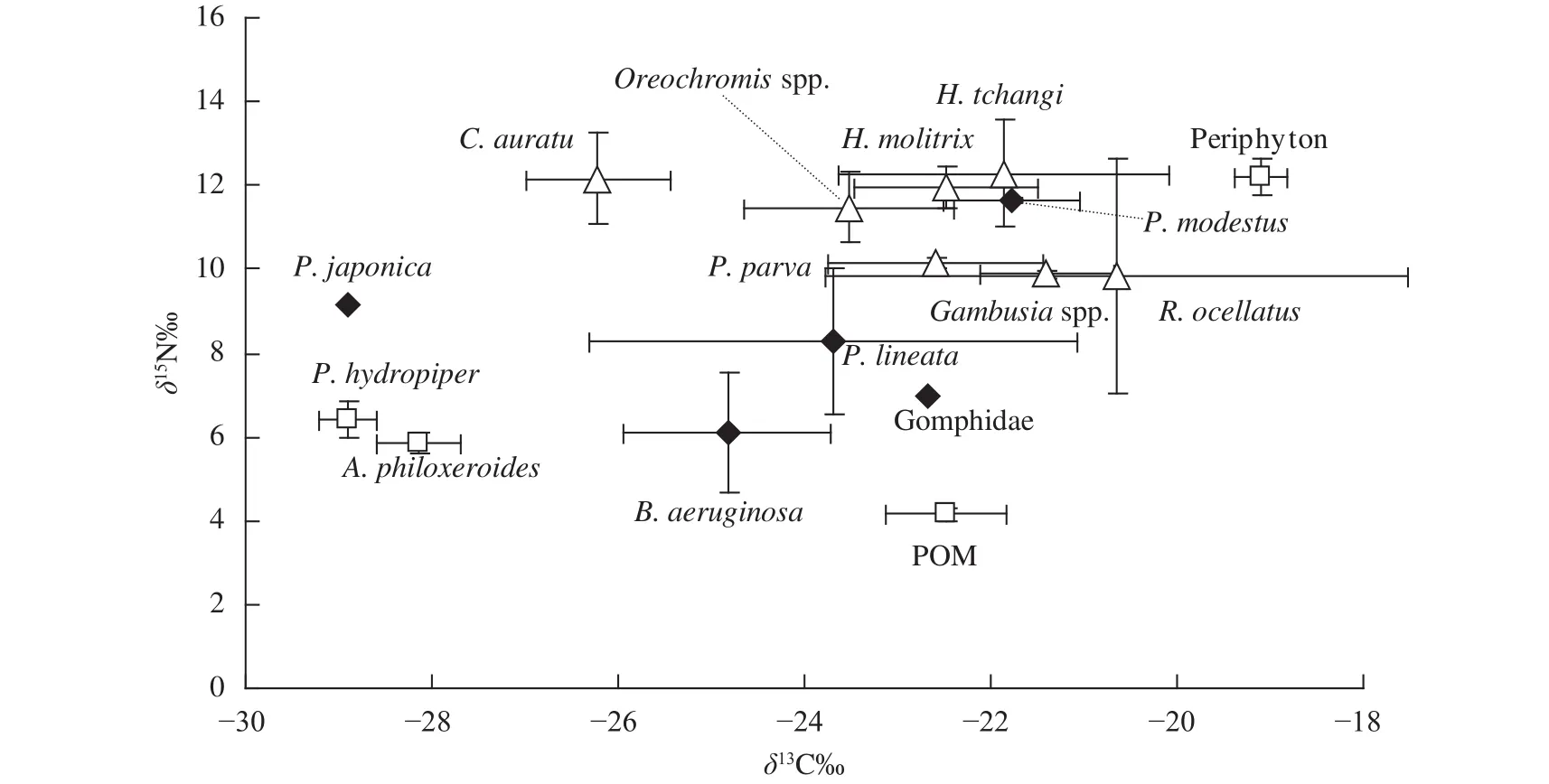
图2 松林水库不同水生生物类群δ13C-δ15N双坐标图Fig. 2 Stable carbon and nitrogen isotope bi-plots based on species collected from the Songlin Reservoir
2.5 消费者生态位空间结构
采用“δ13C–δ15N”组成的二维生态位结合Layman等[19]的同位素营养结构量化指标计算了松林水库消费者的生态位空间结构,并比较了底栖无脊椎动物与鱼类生态位空间结构的异同(表3和图3)。表3显示,底栖无脊椎动物在生态位空间总面积(TA)、营养级长度(NR)、食源多样性(CR)、营养多样性(CD)及物种生态位均匀度(SDNND)等方面都高于鱼类群体,但其营养冗余则相反(NND值越小冗余度越大)。
2.6 Bayesian 混合模型
基于Bayesian混合模型分析了松林水库不同消费者的基础碳来源(表4和图4)。福寿螺和铜锈环棱螺碳源主要来源于POM(30%和27%)、喜旱莲子草(29%和30%)和水蓼(26%和31%);秀丽白虾的碳源以固着藻类(31%)、POM(29%)和喜旱莲子草(21%)为主;POM、固着藻类、喜旱莲子草和水蓼对箭蜓和石蚕的碳源贡献比例较为均衡(表4和图4)。食蚊鱼、麦穗鱼Pseudorasbora parva和鲢Hypophthalmichthys molitrix主要碳源来源于POM,贡献比例平均值分别为37%、43%和31%。上述4种碳源对罗非鱼Oreochromisspp.、黑尾和鲫Carassius auratus贡献比例较为均匀(表4);高体鳑鲏碳源以固着藻类为主(34%)。
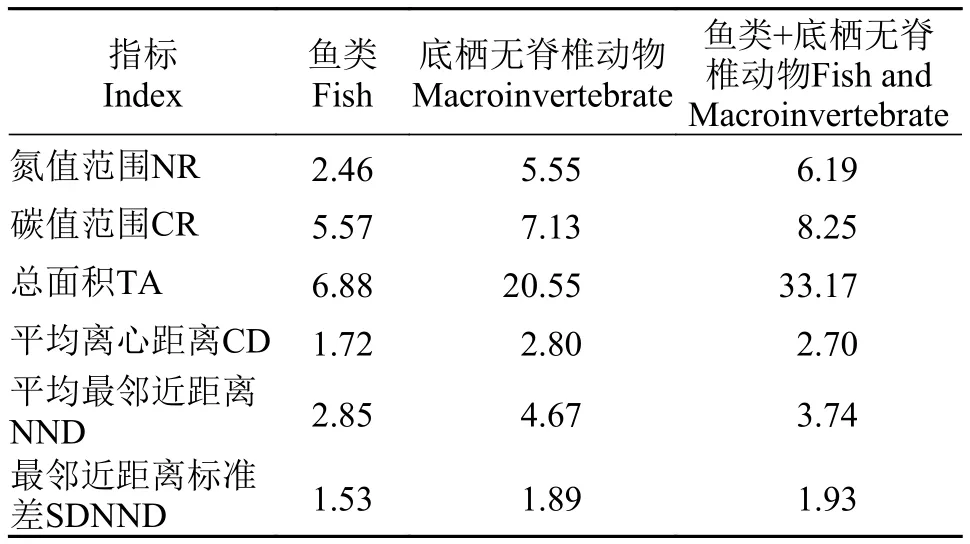
表3 基于Layman形态生态学方法的量化指标Tab. 3 The quantitative index based on the Layman morphological-ecological methodology
3 讨论
3.1 初级生产者同位素特征
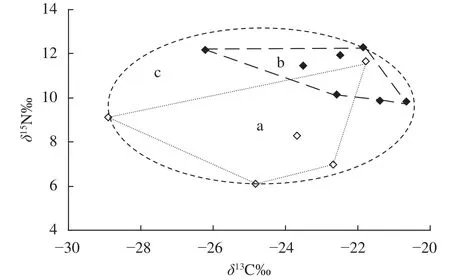
图3 松林水库主要消费者营养结构图Fig. 3 The trophic structure of the main consumers in the Songlin Reservoir
水化学分析显示,采样期间松林水库呈富营养化状态。通常水生高等植物和藻类(浮游藻类和底栖固着藻类)主要通过C3途径利用水体中的CO2合成有机物[20]。Takahashi等[21]认为溶解无机碳浓度是影响δ13C值分馏的主要因素,特别是在夏季富营养化水体中,低浓度的溶解无机碳可引起浮游藻类δ13C值的增加,可能是导致本研究中POM(主要成分为藻类)相对较高的δ13C值的原因,这一现象也与其他水生生态系统研究结果一致[22]。水生维管束植物(水蓼和喜旱莲子草)δ13C值组成恰好介于已报道的湿生植物和陆生植物的范围之间(–23‰— –30‰)[23]。底栖固着藻类的δ13C值显著高于(P<0.05)浮游藻类和水生维管束植物,其原因可能是底层固着藻类在吸收周围环境中的无机碳源时发生的缓慢扩散[24]和富营养化水体底层沉积物碳代谢引起δ13C值增加等因素[25]。研究发现不同初级生产者δ13N值变化明显,特别是底栖固着藻类δ13N值显著高于其他生产者(P<0.05),甚至超过了部分高级捕食鱼类(表1),这可能与松林水库周边大量生活污水和农田径流输入(δ13N值)及生物体自身的生理特征等有关[26]。
3.2 碳源基础与营养结构特征
碳源到消费者的营养转化是驱动食物网能量流动的基础。Bayesian混合模型表明POM(主要成分为藻类)和底栖固着藻类是螺类、秀丽白虾、食蚊鱼、麦穗鱼和鲢等多数消费者的主要基础碳源,该结果与Vander Zanden等[27,28]探究浅水型水库中的研究结论一致;另外,松林水库消费者的C∶N比值(表1)组成也暗示了消费者基础碳源主要来源于内源性营养物质[29]。
食物关系是了解生态系统能量流动的核心,也是群落赖以生存的基础[16]。松林水库食物链长度为3级,鱼类的营养级介于3.09 (高体鳑鲏)—3.82 (黑尾),与Post[16]对淡水生态系统食物网中初级生产者到高级捕食者之间的营养级长度的研究结果吻合,但与我国中部地区的富营养化水体如东湖(4级)[26]、巢湖(4级)[11]相比减少了1个营养等级(最高为3.82)。本研究显示该区域秀丽白虾营养级水平明显高于其他无脊椎动物(P<0.05),可能与其肉食性的摄食生态类型有关[30]。不同鱼类之间δ13C和δ13N值组成无显著差异(P>0.05),且分布较为集中(图2),表明了它们具有相似食物来源。结合同位素数据和相关文献可知,罗非鱼、鲫、麦穗鱼等鱼类食物来源较为广泛,属于典型的杂食性鱼类。此外,松林水库中多数消费者(鱼类和虾)位于食物网中间营养级位置,有利于增加食物网中碳源的流通途径[31],符合能量传递规律[32]。
鱼类物种丰富度与水域面积、生产力之间呈正相关[33]。诸多研究表明亚热带区域的水库或湖泊具有较高的鱼类丰度[34,35]。依据Post[36]提出的插入和补充理论,松林水库(亚热带区域)应该拥有高的鱼类丰富度和较长的食物链,但结果却与之相反。全球范围内随着温度的升高和海拔的降低,淡水生态系统中杂食性鱼类比例呈上升趋势[35];Meerhoff等[34]和Arim等[37]研究发现以杂食性鱼类为优势群体的水域生态系统,因能量限制等因素可导致鱼类物种丰富度和营养级长度下降。因此,杂食性鱼类可能是引起松林水库鱼类多样性和营养级长度下降的重要因素之一。
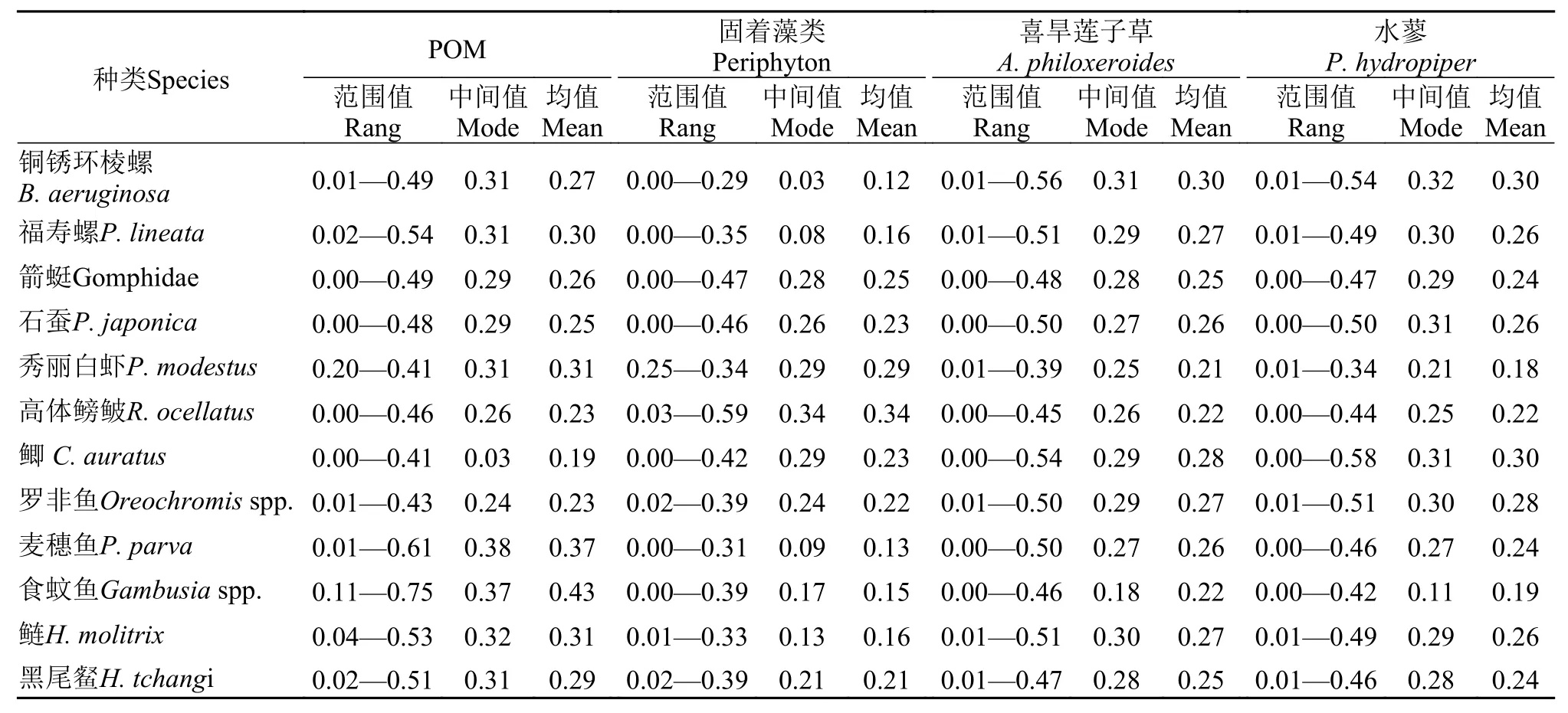
表4 松林水库基础碳源对不同消费者的贡献比例(95%)Tab. 4 Stable isotope analysis in R (SIAR) results of the food sources proportions in the diet of the consumers in the Songlin Reservoir(95%)
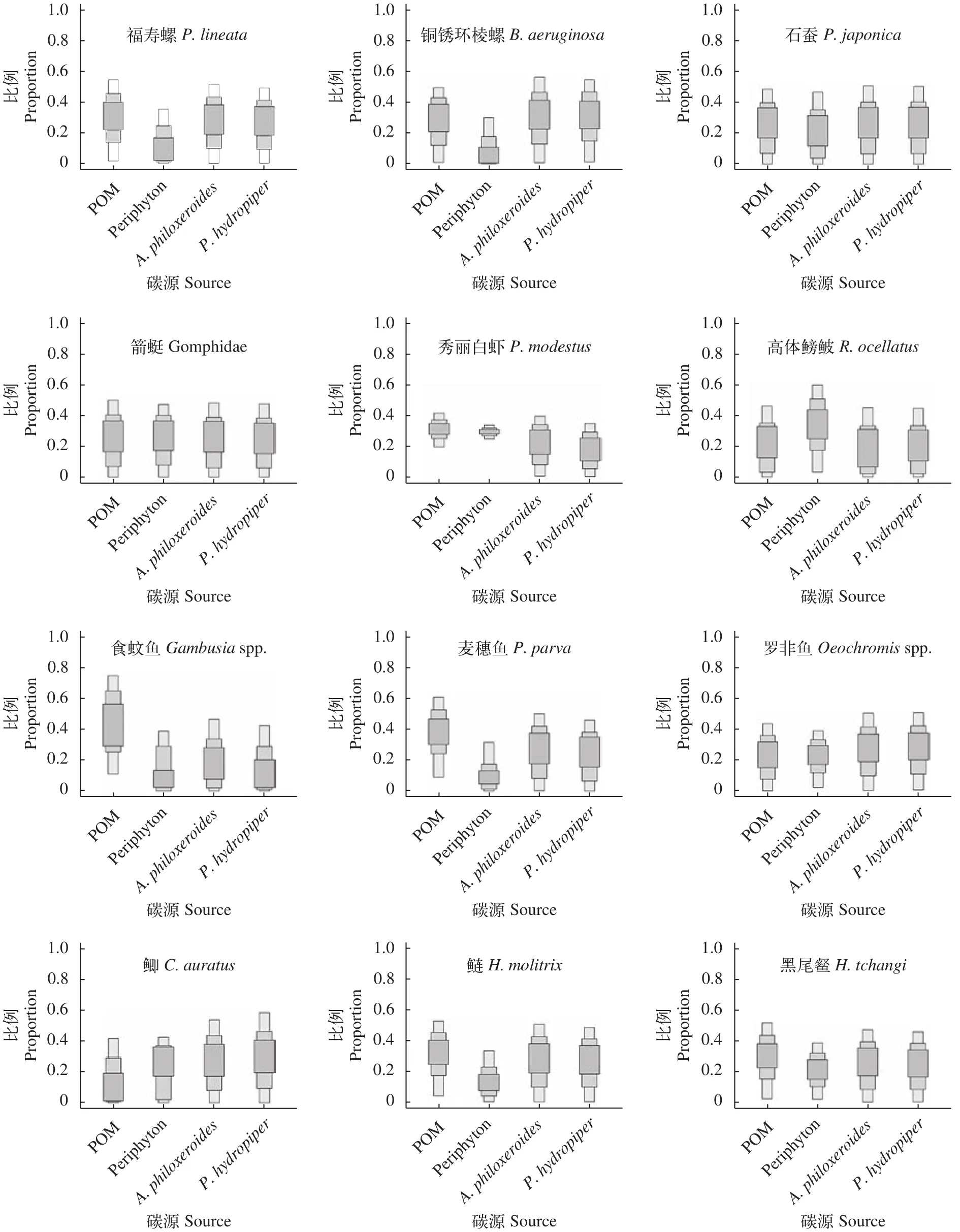
图4 松林水库基础碳源对不同贡献的箱形图Fig. 4 Relative contribution of sources to each consumers in the Songlin Reservoir
3.3 同位素生态位
生物与非生物的稳定同位素比值代表了多维以稳定同位素比值为坐标组成的“δ-空间”被称为同位素生态位(Isotopic niche)[38],反映的是物种的时空整合数据[39],可用于区分食物网中不同营养级位置之间的微小差别[19,40]。本研究中底栖无脊椎动福寿螺(外来物种)和铜锈环棱螺(土著物种)之间存在同位素生态位重叠现象。与底栖动物相比,鱼类群落CR值表明了不同鱼类之间具有明显的同位素生态位重叠现象,特别是外来物种罗非鱼与土著物种鲫、鲢、黑尾之间重叠更为严重(图2)。福寿螺和罗非鱼(外来物种)的CR、NR值大于铜锈环棱螺和鲫,暗示了它们具有广泛的食物来源和较强的环境适应能力,对当地土著物种生存构成一种威胁[29]。本研究发现松林水库NND值较低,杂食性鱼类为优势类群,说明松林水库鱼类群落中存在营养冗余,因此该生态系统抗御外界干扰的抵抗能力较强,生态系统的稳定性也较高[19,32,41]。
致谢:
感谢内江师范学院地理与环境科学学院许斌在计算机分析方面提供的帮助,内江师范学院本科生吴迪、彭敏锐、周鑫、郑武举、刘果等同学在样品采集过程提供的的帮助。
gt;
[1]Constaza R D, Arge R, De Groot R,et al. The value of the world’s ecosystem services and natural capital [J].Nature, 1999, 387(15): 253—260
[2]Ye S W, Yang H B, Chen Y B,et al. Analysis on the development strategies and key techniques of ecological fisheries in the Three Gorges Reservoir, China [J].Acta Hydrobiologica Sinica, 2015, 39(5): 1035—1040 [叶少文, 杨洪斌, 陈永柏, 等. 三峡水库生态渔业发展策略与关键技术研究分析. 水生生物学报, 2015, 39(5):1035—1040]
[3]Vörösmarty C J, McIntyre P B, Gessner M O,et al. Global threats to human water security and river biodiversity[J].Nature, 2010, 467(7315): 555—561
[4]Xie F, Gu J G, Lin Z W. Assessment of aquaticecosystemhealthbasedonprincipal component analysis with entropy weight: A case study of Wanning Reservoir (Hainan Island, China) [J].Chinese Journal of Applied Ecology, 2014, 25(6): 1773—1779 [谢飞, 顾继光, 林彰文.基于主成分分析和熵权的水库生态系统健康评价-以海南省万宁水库为例. 应用生态学报, 2014, 25(6):1773—1779]
[5]Mateo M A, Serrano O, Serrano L,et al. Effects of sample preparation on stable isotope ratios of carbon and nitrogen in marine invertebrates: implications for food空间信息[16,38],web studies using stable isotopes [J].Oecologia, 2008,157(1): 105—115
[6]Fry B, Sherr E B.δ13C Measurements as indicators of carbon flow in marine and freshwater ecosystems [J].Contributions in Marine Science, 1984, 27(8): 13—47
[7]Minagawa M, Wada E. Stepwise enrichment of15N along food chains: further evidence and the relation between15N and animal age [J].Geochimica et Cosmochimica Acta,1984, 48(5): 1135—1140
[8]Hill J M, Jones R W, Hill M P,et al. Comparisons of isotopic niche widths of some invasive and indigenous fauna in a South African river [J].Freshwater Biology, 2015,60(5): 893—902
[9]Wen Z R, Xiong Y, Xu J,et al. The studies on the structures of the food web and the trophic relationships in the Gonghu Bay of the Taihu Lake [J].Acta Hydrobiologica Sinica, 2016, 40(1): 131—138 [温周瑞, 熊鹰, 徐军, 等.太湖贡湖湾食物网特征研究. 水生生物学报, 2016,40(1): 131—138]
[10]Mao Z G, Gu X H, Zeng Q,et al. Food web structure of a shallow ertrophic lake (Lake Taihu, China) assessed by stable isotope analysis [J].Hydrobiologia, 2012, 683(1):173—183
[11]Xu J, Wen Z R, Ke Z X,et al. Contrasting energy pathways at the community level as a consequence of regime shifts [J].Oecologia, 2014, 175(1): 231—241
[12]He J. The design of new building of pine forest in Sonling Reservoir [J].Sichuan Water Conservancy, 2006,27(2): 36—37 [何进. 松林水库新建放空洞设计探讨. 四川水利, 2006, 27(2): 36—37]
[13]State Environmental Protection Administration, the Monitor and Analysis Methods of Water and Wastewater Editorial Committee. The Monitor and Analysis Methods of Water and Wastewater [M]. Beijing: Chinese Environmental Science Press. 2002, 88—284 [国家环境保护总局, 水和废水监测分析方法编委会. 水和废水监测分析方法(第四版). 北京: 中国环境科学出版社. 2002,88—284]
[14]Jin X C, Tu Q Y. Survey Standards of Lake Eutrophication (2nd Edition) [M]. Beijing: China Environmental Science Press.1990, 268—270 [金相灿, 屠清英. 湖泊富营养化调查规范(第二版). 北京: 中国环境科学出版社.1990, 268—270]
[15]Zhang Z S, Huang X F. The Research Methods of Freshwater Plankton [M]. Beijing: Science Press. 1991, 1—414[章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社. 1991, 1—414]
[16]Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions [J].Ecology,2002, 83(3): 703—718
[17]Parnell A C R, Inger S, Bearhip A,et al. Source partitioning suing stable isotopes: coping with too much variation[J].PLoS One, 2010, 5(3): e9672
[18]McCutchan Jr J H W M, Leeis Jr C, Kendall C,et al.Variation in trophic shift for stable isotope ratioss of carbon, nitrogen, and sulfur [J].Oikos, 2003, 102(2):378—390
[19]Layman C A, Arrington D A, Montaña C G,et al. Can stable isotope ratios provide for community-wide measures of trophic structure [J]?Ecology, 2008, 89(8):2358—2367
[20]Laws E A, Popp B N, Bidigare R R,et al. Dependance of phytoplankton carbon isotope composition on growth rate and [CO2] aq: theoretical considerations and experimental results [J].Geochimica Et Cosmochimica Acta, 1995,59(6): 1131—1138
[21]Takahashi K, Yoshioka T, Wada E,et al. Temporal variations in carbon isotope ratio of phytoplankton in a eutrophic lake [J].Journal of Plankton Research, 1990,12(4): 799—808
[22]Zhang H, Peng L, Gu B,et al. Effect of trans-reservoir water supply on carbon and nitrogen stable isotope composition in hydrologically connected reservoirs in China[J].Chinese Journal of Oceanology amp; Limnology, 2016:1—10
[23]Lu F Y, Liu Z Q, Ji H B. Carbon and nitrogen isotopes analysis and sources of organic matter in the upper reaches of the Chaobai River near Beijing [J].China Science China Earth Sciences, 2013, 56(2): 217—227
[24]Yoshioka T, Wada E, Yatsuka S. Isotopic characterization of Lake Kizaki and Lake Suwa [J].Japanese Journal of Limnology, 1988, 49(2): 119—128
[25]Kendall C, Silva S R, Kelly V J. Carbon and nitrogen isotopic compositions of particulate organic matter in four large river systems across the United States [J].Hydrological Processes, 2001, 15(7): 1301—1346
[26]Xu J, Xie P, Zhang M,et al. Variation in stable isotope signatures of seston and a zooplanktivorous fish in a eutrophic Chinese lake [J].Hydrobiologia, 2005, 541(1):215—220
[27]Vander Zanden M J, Fetzer W W. Global patterns of aquatic food chain length [J].Oikos, 2007, 116(8):1378—1388
[28]Vander Zanden M J, Vadeboncoeur Y, Chandra S. Fish reliance on littoral-benthic resources and the distribution of primary production in lakes [J].Ecosystems, 2011,14(6): 894—903
[29]Dalu T, Weyl Olaf L F, Froneman P W,et al. Trophic interactions in an austral temperate ephemeral pond inferred using stable isotope analysis [J].Hydrobiologia,2016, 768(1): 81—94
[30]Zhang H, Xie P, Wu G G,et al. Studies on trophic niches ofMacrobrachium nipponensisandExopalaemon modestus[J].Research of Environmental Sciences, 2013,26(1): 22—26 [张欢, 谢平, 吴功果, 等. 日本沼虾与秀丽白虾的营养生态位. 环境科学研究, 2013, 26(1):22—26]
[31]Post D M, Takimoto G. Proximate structural mechanismsfor variation in food-chain length [J].Oikos, 2007,116(5): 775—782
[32]Iglesias C, Meerhoff M, Johansson L S,et al. Stable isotope analysis confirms substantial differences between subtropical and temperate shallow lake food webs [J].Hydrobiologia, 2017, 784(1): 111—123
[33]DodsonS I, Arnott S E, Cottingham K L. The relationship in lake communities between primary pro-ductivity and species richness [J].Ecology, 2000, 81(10): 2662—2679
[34]Meerhoff M, Clemente J M, Mello F T D,et al. Can warm climate-related structure of littoral predator assemblies weaken the clear water state in shallow lakes [J]?Global Change Biology, 2007, 13(9): 1888—1897
[35]González-Bergonzoni I, Meerhoff M, DavidsonT,et al.Meta-analysis shows a consistent and strong latitudinal pattern in fish omnivory across ecosystems [J].Ecosystems, 2012, 15(3): 492—503
[36]Post D M. The long and short of food-chain length [J].Trends in Ecology and Evolution, 2002, 17(6): 269—277
[37]Arim M, Marquet P A. On the relationship between trophic position, body mass and temperature: reformulating the energy limitation hypothesis [J].Oikos, 2007, 116(9):1524—1530
[38]West J B, Bowen G J, Cerling T E,et al. Stable isotopes as one of nature’s ecological recorders [J].Trends in Ecology and Evolution, 2006, 21(7): 408—414
[39]Newsome S D, Martinez del Rio C, Bearhop S,et al. A niche for isotopic ecology [J].Frontiers in Ecology and the Environment, 2007, 5(8): 429—436
[40]Bearhop S, Adams C E, Waldron S,et al. Determining trophic niche width: a novel approach using stable isotope analysis [J].Journal of Animal Ecology, 2004, 73(5):1007—1012
[41]Baiser B, Ardeshiri R S, Ellison A M. Species richness and trophic diversity increase decomposition in a coevolved food web [J].PLoS One, 2011, 6(5): e20672
CHARACTERIZING THE TROPHIC STRUCTURE OF A SMALL RESERVOIR IN SOUTHEAST OF SICHUAN PROVINCE USING STABLE CARBON AND NITROGEN ISOTOPE ANALYSIS
LI Bin1,2, WANG Zhi-Jian2, XU Dan-Dan1, WANG Yong-Min1and TAO Min1
(1. School of Life Sciences, Neijiang Normal University, Neijiang 641100, China; 2. Key Laboratory of Freshwater Fish Reproduction and Development (Ministry of Education), Southwest University, Chongqing 400715, China)
Reservoir plays important roles in realizing the sustainable development of economy and society. It can provide water resource to human beings for living, industrial and agricultural usage, and for flood control, power generation and regional ecological service as well. The sustainable utilization of the reservoir water resource is the prerequisite for fishery industry development. However, following with the impacts of increasing human activities, pollutant input and water eutrophication, the ecological environment gets deteriorated overtime, which was revealed by water balance disorders, such as sharp decrease of biodiversity and others. In order to better understand the trophic structure characteristics of reservoir ecosystem in China, Songlin, a typical small reservoir in Southeast Sichuan province, was selected for the stable isotope analysis. The basic carbon sources of different consumers were analysed based on the Bayesian mix-model (SIAR), and six nutrition structure quantitative indicators inδ13C—δ15N isotopic niches were calculated as well. Results showed a eutrophication state of this small reservoir, and theδ13C andδ15N values of primary producers such as POM (mainly the planktonic algae), periphyton,Alternanthera philoxeroidesandPolygonum hydropiperwere within the scopes of –29.20‰—18.81‰ and 4.01‰—12.73‰, respectively. POM and periphyton were found as the main carbon sources for most consumers. The food web trophic level length of Songlin reservoir was calculated at Level 3, with omnivorous fishes as superior group and existed the nutrition redundancy phenomenon, which suggested a relatively stable fish community structure in Songlin reservoir. Additionally, niche overlaps and competitions were found between invasive species (such asPomacea lineataandOreochromissp) and native species (such asBellamya aeruginosaandCarassius auratus, etc). Taking together, we suggested to enhance the management of aquatic organism resources, and powerful strategies must be taken to prevent and control the invasive alien pest for better protection on the biodiversity of Sonlin reservoir.
Reservoir; Trophic structure;δ13C andδ15N; Isotope niche; Bayesian mixing-model
Q148
A
1000-3207(2017)06-1345-09
2016-12-14;
2017-04-10
四川省教育厅重点项目(14ZA0251); 三峡库区小江汉丰湖流域水环境综合防治与示范课题(2013ZX07104-004-005)资助[Supported by the Key Fund Project of Sichuan Provincial Department of Education (14ZA0251); The Demonstration Project and Water Environmental Prevention Planning of Hanfeng Lake Reaches in Three Gorge Reservoir (2013ZX07104-004-005)]
李斌(1981—), 男, 河南信阳人; 博士, 副教授; 主要从事水域生态学研究。E-mail: libiin2004xinyang@126.com
陶敏, E-mail: celery_319_07@126.com
10.7541/2017.166
