翘嘴红鲌雌雄基因组差异及太湖野生群体遗传多样性现状的AFLP分析
2017-11-29刘祥芳郑建波贾永义顾志敏
刘祥芳 郑建波 贾永义 顾志敏 罗 琛
(1. 浙江大学生命科学学院, 杭州 310058; 2. 浙江省淡水水产研究所, 湖州 313001)
翘嘴红鲌雌雄基因组差异及太湖野生群体遗传多样性现状的AFLP分析
刘祥芳1郑建波1贾永义2顾志敏2罗 琛1
(1. 浙江大学生命科学学院, 杭州 310058; 2. 浙江省淡水水产研究所, 湖州 313001)
为发展翘嘴红鲌Erythroculter ilishaeformis(Bleeker)性别控制育种和单性养殖技术, 利用AFLP技术对翘嘴红鲌太湖野生群体中雌、雄各20个个体的基因组进行了遗传多样性和雌雄差异分析。用筛选出的8对AFLP多态性引物在太湖野生群体基因组中共检测到319个位点, 其中多态性位点185个。通过对所有多态性位点在个体中的分布进行分析, 发现一个只在雄性个体中稳定存在、在雌性个体中缺失的差异位点。在人工雌核发育翘嘴红鲌群体只有雌性个体, 其基因组中也检测不到该差异位点的存在。这些结果表明翘嘴红鲌性别分化可能受到严格的遗传调控, 该差异位点是雄性特有的性别分子标记。翘嘴红鲌太湖野生群体目前的多态位点比率P=57.99%, 观测等位基因数Na=1.5799±0.4943, 有效等位基因数Ne=1.3859±0.3971, Shannon’s信息指数I=0.3221±0.2987。这些统计分析结果与10年前对该群体的AFLP研究结果(P=51.21%,Na=1.512±0.500,Ne=1.252±0.371,I=0.218±0.275)相比没有显著差别(P>0.05), 说明该群体目前还具有适度的遗传多样性。
翘嘴红鲌; 基因组分析; 雌雄差异; 遗传多样性
翘嘴红鲌Erythroculter ilishaeformis(Bleeker),又名翘嘴鲌, 隶属鲤科、鲌亚科、鲌属, 为中上层大型广温性淡水鱼类, 是我国分布最广的经济鱼类之一。太湖流域的翘嘴红鲌品质优良, 与银鱼、白虾并称“太湖三白”而名闻天下。近年来, 随着翘嘴红鲌人工养殖、繁殖技术的成熟及推广, 该鱼已经成为中国特别是长江三角洲地区的主要名特淡水养殖品种[1]。翘嘴红鲌雌鱼比雄鱼的个体大、生长速度快, 是有性别异形[2]特征的鱼类。已有的育种实践表明开发性别特异性分子标记, 是发展性别控制育种技术, 实现这些鱼类单性养殖而提高其养殖经济效益的重要技术途径[3]。但目前与翘嘴红鲌性别决定和分化相关的关键调控基因以及调控机制都不清楚, 与性别相关的基因组分子标记未见报道,限制了性别控制育种和单性养殖技术在翘嘴红鲌养殖业中的应用。
扩增片段长度多态性(Amplified fragment length polymorphism, AFLP)技术是1993年由Vos等[4]发明的一种DNA指纹技术。AFLP结合了限制性内切酶片段长度多态性(Restriction fragment length polymorphism, RFLP)技术的稳定性与PCR技术的高效性, 省去了RFLP的繁杂过程, 同时又克服了随机扩增多态性DNA (Random amplified polymorphic DNA, RAPD)技术不稳定的缺点, 是一种能有效揭示生物种间和种内遗传差异的高容量标记技术[5,6]。AFLP筛选信息量很大、可重复性很高[4], 这些优点在性别连锁标记的筛选中是微卫星(Microsatellite)、RAPD等其他遗传标记无可比拟的[7], 已成功应用于多种动植物高密度遗传图谱的构建、分子标记的筛选以及遗传多样性的研究[8—13]。目前, 利用AFLP技术已筛选到尼罗罗非鱼(Oreochromis niloticus)[14]和黄颡鱼(Pelteobagrus fulvidraco)[11,12]等重要水产养殖鱼类的雄性或雌性特异分子标记并成功转化为SCAR (Sequence characterized amplified regions)标记。黄颡鱼的性别分子标记已经成功应用于性别控制育种和单性养殖[2,11]。因此, 本研究运用AFLP技术对翘嘴红鲌太湖野生群体中大量雌、雄个体的基因组进行了遗传多样性分析, 发现和确定了翘嘴红鲌基因组DNA中与性别相关的分子标记。这些结果为深入研究翘嘴红鲌性别决定的遗传机制[13]、发展翘嘴红鲌性别控制育种和单性养殖技术提供了基础, 也为了解翘嘴红鲌太湖野生群体目前的遗传多样性状况提供了有重要价值的资料。
1 材料与方法
1.1 材料和试剂
从浙江湖州太湖水域多个地点随机选取野生翘嘴红鲌雌、雄各20尾, 剪取尾鳍置于液氮中进行快速冷冻处理, 随后置于–80℃冰箱中保存。实验中所用引物皆由上海生工生物工程有限公司合成,酶切相关试剂购自Thermo Scientific和BioLabs公司, 连接、预扩增及选择性扩增所用相关试剂均购自TaKaRa公司。
1.2 DNA的提取
参照改进的酚/氯仿法[15]提取基因组DNA。提取的基因组DNA用分光光度计检测OD值并进行琼脂糖凝胶电泳检查。A260nm/A280nm比值在1.7—2.0,且电泳结果显示无降解、无蛋白质污染的基因组DNA用于后续的AFLP分析。
1.3 AFLP分析
AFLP反应体系参照Vos等[4]描述的方法进行,并对其中影响实验结果的关键因素进行了优化。AFLP接头及预扩增引物序列见表1。正向和反向各8条选择性扩增引物序列见表2。由这些正、反向的选择性扩增引物排列组合成共64对不同的选择性扩增引物。
限制性酶切使用EcoRⅠ(Thermo Scientific)和MseⅠ(Bio Labs)两种限制性内切酶, 在37℃水浴中同时对300 ng基因组DNA进行酶切, 反应4h后于75℃金属浴中对限制性内切酶灭活12min。随后取少量酶切产物在1%(w/v)的琼脂糖凝胶中进行电泳检测, 以确保基因组DNA被充分酶切。
连接取10 μL酶切产物, 使用1 μLMseⅠ接头(50 μmol/ L)和1 μLEcoRⅠ接头(5 μmol/ L)于22℃水浴中连接过夜。
预扩增预扩增反应的体系为: 连接产物3 μL,MseⅠ预扩增引物(Mo 50 ng/μL) 1 μL,EcoRⅠ预扩增引物(Eo 50 ng/μL) 1 μL, PCR Buffer 2 μL, rTaq酶1 U, dNTP Mix (10 mmol/L)2 μL及ddH2O 10.8 μL。PCR反应条件为: 先于94℃预处理5min; 而后按照94℃ 20s, 56℃ 30s, 72℃ 2min, 循环20次。预扩增完成后, 用ddH2O将产物稀释20倍,–20℃条件下保存。
选择性扩增取1 μL稀释的预扩增产物, 加入14 μL如下反应液:EcoRⅠ引物(2.5 μm/L) 1 μL,MseⅠ引物(5 μm/L) 1 μL, dNTP Mix (10 mmol/L)1.2 μL, rTaq酶1 U, PCR Buffer 1.5 μL, ddH2O 9.1 μL,混合后按如下PCR程序扩增: 94℃ 5min; 94℃ 30s,65℃ 30s (每循环降低 0.7℃), 72℃ 60s, 循环14次;94℃ 30s, 56℃ 30s, 72℃ 60s, 循环23次。
聚丙烯酰胺凝胶电泳及银染选择性扩增产物经8%聚丙烯酰胺凝胶电泳分离, 随后银染显色, 银染完成后用凝胶成像系统扫描, 保存扫描结果用于后续的数据统计与分析。
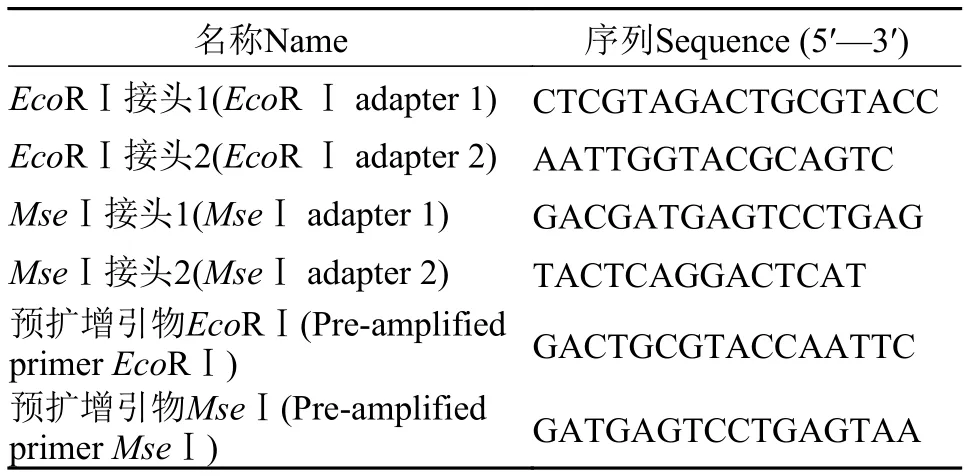
表1 AFLP接头及预扩增引物序列Tab. 1 Sequences of adapters and pre-amplified primers in AFLP analysis

表2 用于AFLP分析的选择性扩增引物的名称及序列Tab. 2 Selective amplification primers and their sequences used in AFLP analysis
1.4 数据统计与分析
AFLP标记为显性标记, 在后代中的遗传和分离中符合Mendel式遗传规律, 种群中的AFLP标记位点遵循Hardy-Weinberg平衡。首先将重复性好且清晰的AFLP条带进行统计, 建立原始数据矩阵。然后利用Popgen32软件统计位点总数和多态位点数, 并计算出多态位点比率(Percentage of polymorphic loci-P)、观测等位基因数(Observed number of alleles-Na)、有效等位基因数(Effective number of alleles-Ne)以及Nei’s多样性指数(Nei’s diversity index-H)和Shannon’s信息指数(Shannon’s information index-I)。
(1)多态位点比率是指在所有的检测到的位点中多态位点所占的比例, 用其作为度量遗传变异水平高低的指标。一个种群多态位点比率高, 说明这个种群适应环境的能力强, 遗传变异水平高; 反之,一个种群多态位点比率低, 说明这个种群适应能力弱, 遗传变异水平低。
计算公式为:P(%)=多态位点数/位点总数×100
(2)观测等位基因数是所检测到的等位基因数与所检测到的总位点数之比。
计算公式为:Na=(多态位点数×2+单态位点数)/位点总数。
(3)有效等位基因数是纯合度的倒数, 反映了群体的变异程度。有效等位基因数越接近观测等位基因数, 说明等位基因在群体的分布越均匀。
(4)Nei’s多样性指数为各个位点的平均杂合度,可以反映各群体在几个位点上的遗传差异, 一般认为它是度量群体遗传变异的一个较适合的参数。
H的计算公式为:H=1–∑Pi2,Pi为单个位点上的等位基因频率。
(5)Shannon’s信息指数用来衡量群体多样性的高低, 其值越大, 说明群体内的变异越大。
I=–∑XiLnXi/N,Xi表示位点i在某个群体中的出现频率,N表示该群体中检测到的位点总数。
最后, 利用SPSS 19.0中的t检验对翘嘴红鲌太湖野生群体2007年与2016年的遗传参数进行差异显著性检验,P值大于0.05认为差异不显著。
2 结果
2.1 多态性AFLP引物的筛选
为从64对选择性扩增引物组合中筛选出多态性引物, 我们在随机取样的6条翘嘴红鲌基因组样本上用这些引物进行了PCR产物扩增和电泳分析。结果表明有15对引物组合没有扩增出清晰稳定的产物电泳条带, 49对引物组合能扩增出清晰稳定的产物电泳条带。对电泳产物条带的分析显示有42对引物在不同个体样品上扩增出了不同的产物, 为多态性引物; 有7对引物在所有个体样品上都扩增出同样的产物, 为单态性引物(表3)。
然后我们从42对多态性引物中选择图谱清晰、重复性好且扩增产物条带数较多、复杂程度中等的8对引物组合(E1/M7、E2/M5、E4/M5、E5/M1、E6/M6、E7/M5、E8/M6和E8/M8), 作为对翘嘴红鲌太湖野生群体的所有个体基因组进行扩增分析的引物。
2.2 翘嘴红鲌太湖流域群体遗传多样性现状
用所筛选的8对多态性引物, 对翘嘴红鲌太湖野生群体40尾(雌、雄各20尾)个体的基因组分别进行PCR扩增和电泳的分析统计结果见表4。统计时取长度在80—587 bp, 且清晰、稳定的电泳条带(图1)。8对多态性引物扩增出的位点数从35到45不等,扩增出的多态位点数从18到31不等。多态位点比率最高的引物组合是E8/M6(73.81%), 最低的引物组合是E4/M5(46.15%)。8对多态性引物总共扩增出319个可清楚分辨的位点, 其中多态性位点185个。遗传多样性统计分析结果显示太湖流域翘嘴红鲌野生群体目前的多态位点比率为57.99%; 观测等位基因数为1.5799±0.4943; 有效等位基因数为1.3859±0.3971; Nei’s多样性指数为0.2191±0.2102;Shannon’s信息指数为0.3221±0.2987(表5)。

表3 选择性扩增引物筛选结果Tab. 3 Results of selective amplification primers screening results
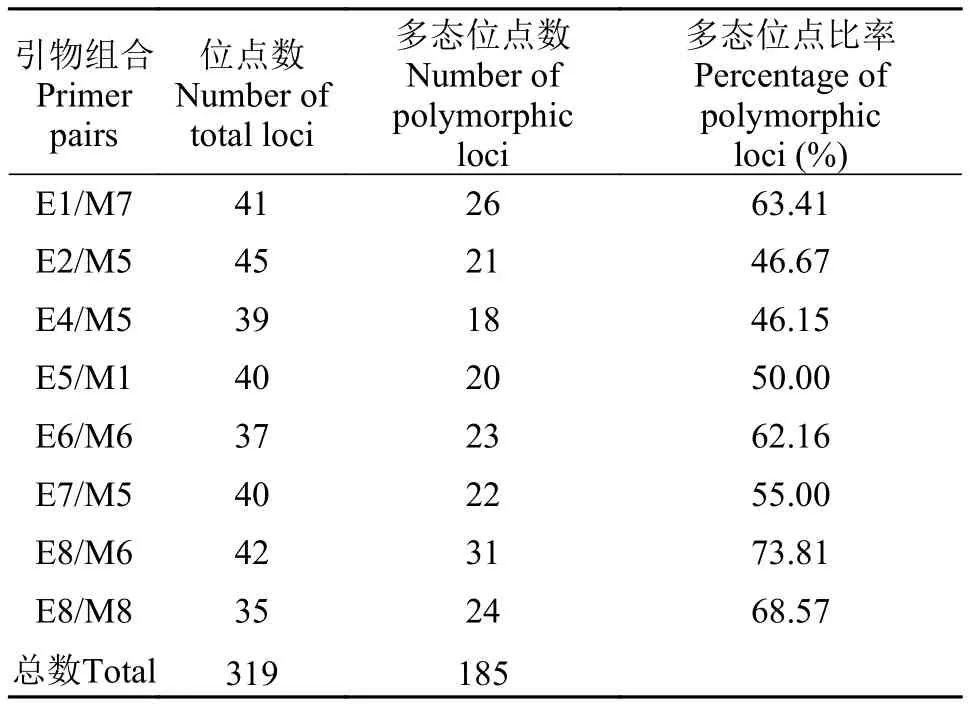
表4 八对选择性扩增引物组合扩增出的位点数和多态位点数Tab. 4 Number of total loci and polymorphic loci amplified by 8 primer pairs
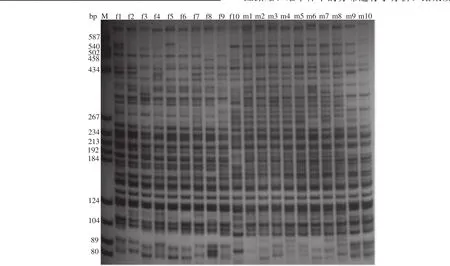
图1 引物E2/M5扩增产物的电泳图谱Fig. 1 Gel electrophoretogram of the PCR products amplified by primer pair of E2/M5
太湖流域翘嘴红鲌群体的遗传多样性已有研究报道, 2007年, 李建林等[16]采用RAPD技术, 王伟等[17]采用AFLP和ISSR (Inter-simple sequence repeat)技术分别对太湖野生和养殖群体的遗传多样性进行了分析。为了解太湖流域翘嘴红鲌野生群体的遗传多样性10年来的变化状况, 我们将目前的遗传多样性状态与2007年王伟[17]用AFLP方法所得的结果进行了比较(表5)。分析结果表明翘嘴红鲌太湖野生群体的遗传多样性水平与2007年王伟[17]的AFLP研究结果差异不显著(P>0.05)。这些结果说明翘嘴红鲌太湖野生群体的遗传多样性仍与2007年一样处在较高的水平, 没有降低。
2.3 翘嘴红鲌性别差异的AFLP标记
我们进一步对检测到的所有多态位点在翘嘴红鲌雌、雄个体中的分布进行了分析。结果发现在引物组合E8/M6所检测到的多态性位点中有一个差异位点在雄性个体中稳定存在, 在雌性个体中不存在(图2A)。这一结果提示该位点可能是翘嘴红鲌雄性基因组中特有的位点。为了确定该位点与翘嘴红鲌性别的关联性, 我们检测了人工诱导雌核发育群体的性别。在所检测的2个不同雌核发育群体的483条鱼和832条鱼中, 没有检测到雄性个体的存在。我们进一步用E8/M6引物组合对随机选取的20尾第二代雌核发育翘嘴红鲌个体的基因组进行了分析。结果表明野生群体中存在的很多等位基因在雌核发育群体中已经丢失, 其电泳条带数远少于野生群体, 不同的个体基因型差异很小, 没有在任何个体电泳图谱的相应位置检测到可分辨的雄性特有电泳条带的存在(图2B)。
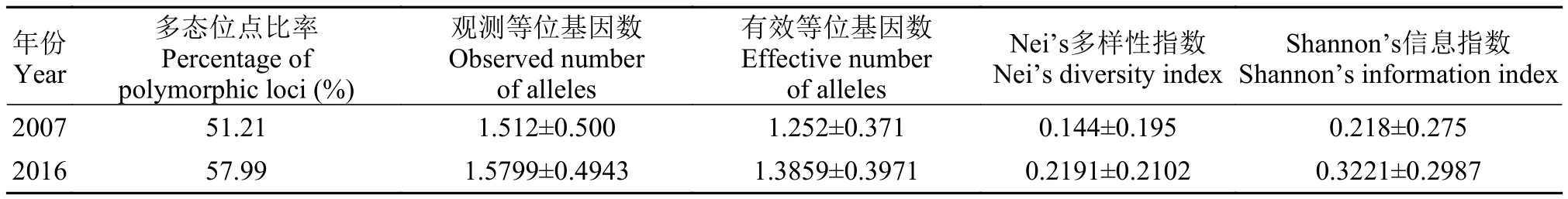
表5 翘嘴红鲌太湖野生群体2007年与2016年的遗传参数比较(平均值±标准误)Tab. 5 Comparison of genetic diversity parameters for E. ilishaeformis (Mean±SE)
3 讨论
为研究翘嘴红鲌雌、雄个体的遗传差异以及其太湖流域野生群体的遗传多样性现状, 我们对翘嘴红鲌太湖野生群体雌、雄各20尾个体的基因组进行了AFLP分析。实验筛选出8对高效的AFLP多态性引物, 用这些引物从翘嘴红鲌太湖野生群体大量雌、雄个体的基因组检测到了185个多态性位点。通过分析这些多态性位点在雌、雄个体中的分布, 我们发现一个在雄性个体中稳定存在、在雌性个体中不存在的差异位点。在各有数个体的2个不同翘嘴红鲌人工雌核发育群体中都没有检测到雄性个体的存在, 也没有在其基因组中检测到该差异位点的存在。这些结果说明所检测到的性别差异位点是翘嘴红鲌雄性特有的性别分子标记; 翘嘴红鲌可能出现了初步的性染色体分化, 其性别分化受到了严格的遗传调控。因此, 通过人工诱导雌核发育获得全雌性鱼, 再对部分雌核发育鱼进行人工诱导性逆转获得生理上为雄性而遗传上为雌性的个体即可进行性别控制育种, 实现翘嘴红鲌的全雌养殖。
雄性特异AFLP分子标记的获得, 为深入研究翘嘴红鲌性别决定的遗传机制、发展翘嘴红鲌的性别控制育种和单性养殖技术提供了基础。目前我们克隆了该雄性特异AFLP分子标记并进行了测序分析, 确定了该序列为非编码序列, 并正在对其连锁基因进行分析, 以将其转换为SCAR标记。
翘嘴红鲌太湖野生群体目前的多态位点比率P、观测等位基因数Na、有效等位基因数Ne以及Shannon’s信息指数I与王伟[17]在2007年对太湖野生群体遗传多样性进行的AFLP分析数据差异不显著(P>0.05), 说明该群体目前还具有适度的遗传多样性。太湖是中国五大淡水湖之一, 河港纵横, 河口众多, 有主要进出河流50余条, 不同群体间基因交流机会大, 良好的自然条件为翘嘴红鲌提供了优越的生活环境, 可能是其遗传多样性仍然能保持在较高水平的原因。但是过度捕捞、环境污染以及人工养殖的单一生态环境和高近亲繁殖机率仍然是翘嘴红鲌太湖野生群体遗传多样性的重要威胁, 在渔业管理中不能过度乐观。

图2 引物E8/M6扩增产物的电泳图谱(箭头示差异条带)Fig. 2 Gel electrophoretogram of the PCR products amplified by primer pair of E8/M6(arrows indicate the differential band)
[1]Gu Z M, Zhu J J, Jia Y Y,et al. Research on embryonic and postembryonic development ofErythroculter ilishaeformisBleeker of Taihu Lake [J].Journal of Fishery Sciences of China, 2008, 15(2): 204—214 [顾志敏, 朱俊杰,贾永义, 等. 太湖翘嘴红鲌胚胎发育及胚后发育观察.中国水产科学, 2008, 15(2): 204—214]
[2]Mei J, Gui J F. Genetic basis and biotechnological manipulation of sexual dimorphism and sex determination in fish(review) [J].Science China Life Sciences, 2014, 44(12):1198—1212 [梅洁, 桂建芳. 鱼类性别异形和性别决定的遗传基础及其生物技术操控. 中国科学: 生命科学,2014, 44(12): 1198—1212]
[3]Gui J F, Zhu Z Y. Molecular basis and genetic improvement of economically important traits in aquaculture animals [J].Science Bulletin, 2012, 57(15): 1751—1760
[4]Vos P, Hogers R, Bleeker M,et al. AFLP: a new technique for DNA fingerprinting [J].Nucleic Acids Research, 1995, 23(21): 4407—4414
[5]Chen J, Li Q, Wang G Y,et al. Analysis of the hybrid F1betweenerythroculter ilishaeformis♀×Ancherytherythroculter nigrocauda ♂by AFLP marker technique [J].Acta Hydrobiologica Sinica, 2014, 38(5): 891—896 [陈见, 李清, 王贵英, 等. 翘嘴红鲌(♀)×黑尾近红鲌(♂)杂种F1的AFLP分析. 水生生物学报, 2014, 38(5): 891—896]
[6]Zhang J B, Huang L M. Analysis of genetic diversity ofLutganus argentimaculatusby AFLP molecular technique [J].Journal of Tropical Oceanography, 2004,23(5): 50—55 [张俊彬, 黄良民. 紫红笛鲷遗传多样性的AFLP分析. 热带海洋学报, 2004, 23(5): 50—55]
[7]Griffiths R, Orr K. The use of amplified fragment length polymorphism (AFLP) in the isolation of sex-specific markers [J].Molecular Ecology, 1999, 8(4): 671—674
[8]Herbergs J, Siwek M, Crooijmans R P,et al. Multicolour fluorescent detection and mapping of AFLP markers in chicken (Gallus domesticus) [J].Animal Genetics, 1999,30(4): 274—85
[9]Ajmone-Marsan P, Vecchiotti-Antaldi G, Bertoni G,et al.AFLP™markers for DNA fingerprinting in cattle [J].Animal Genetics, 1997, 28(6): 418—426
[10]Otsen M, Den B M, Kuiper M T,et al. Use of AFLP markers for gene mapping and QTL detection in the rat[J].Genomics, 1996, 37(3): 289—294
[11]Wang D, Mao H L, Chen H X,et al. Isolation of Y- and X-linked SCAR markers in yellow catfish and application in the production of all-male populations [J].Animal Genetics, 2009, 40(6): 978—981
[12]Dan C, Mei J, Wang D,et al. Genetic differentiation and efficient sex-specific marker development of a pair of Y-and X-linked markers in yellow catfish [J].International Journal of Biological Sciences, 2013, 9(10): 1043—1049
[13]Pan Z J, Li X Y, Zhou F J,et al. Identification of sex-specific markers reveals male heterogametic sex determination inPseudobagrus ussuriensis[J].Marine Biotechnology, 2015, 17(4): 441—451
[14]Sun Y L, Jiang D N, Zeng S,et al. Screening and characterization of sex-linked DNA markers and marker-assisted selection in the Nile tilapia (Oreochromis niloticus)[J].Aquaculture, 2014, 433(5): 19—27
[15]Lin K D, Luo C. Preliminary study on applicability of microsatellite primers developed from common carp for genomic analysis of grass carp [J].Acta Laser Biology Sinica, 2003, 12(2): 121—127 [林凯东, 罗琛. 鲤的微卫星引物对草鱼基因组分析适用性的初步研究. 激光生物学报, 2003, 12(2): 121—127]
[16]Li J L, Wu T T. Analysis of the genetic diversity in Suzhou and Yixing cultivated populations of Topmouth Culter [J].Biotechnology, 2007, 17(2): 26—29 [李建林, 吴婷婷. 翘嘴红鲌苏州和宜兴养殖种群的遗传多样性分析. 生物技术, 2007, 17(2): 26—29]
[17]Wang W. Study on population genetic diversity ofCulter alburnusand phylogeny of Cultrinae [D]. PhD thesis.East China Normal University, Shanghai. 2007 [王伟. 翘嘴鲌(Culter alburnus)群体遗传多样性及鲌亚科鱼类系统发生的研究. 博士学位论文, 华东师范大学, 上海.2007]
AFLP ANALYSIS ON GENOMIC SEXUAL DIMORPHISM OF ERYTHROCULTER ILISHAEFORMIS AND CURRENT GENETIC DIVERSITY OF THE TAIHU STRAIN
LIU Xiang-Fang1, ZHENG Jian-Bo1, JIA Yong-Yi2, GU Zhi-Min2and LUO Chen1
(1. College of Life Sciences, Zhejiang University, Hangzhou 310058, China;2. Zhejiang Institute of Freshwater Fisheries, Huzhou 313001, China)
The Taihu strain ofErythroculter ilishaeformisis a premium aquaculture species and is now widely cultivated in the Yangtze River Delta freshwater region. Under captivity, sexual dimorphism in somatic growth is very common with female being faster grower and having larger body size compared with male. Therefore, manipulating sex for all-female population ofE. ilishaeformiscan substantially increase aquaculture production and economic benefits.However, the key regulatory genes and mechanisms affecting sex determination and differentiation ofE. ilishaeformisare still unclear and no gender-related molecular marker has been reported yet. Consequently, developing the technique for the monosexual production ofE. ilishaeformisremains difficult in the breeding program at present. To develop the technology, the genetic diversity and genomic sexual dimorphism of the Taihu strain were analyzed by AFLP (amplified fragment length polymorphism) molecular marker technique. The sampling size was 20 male and 20 female individuals, respectively. A total of 319 loci were detected using 8 pairs of polymorphic primers. Of the 319 detected loci,185 loci displayed polymorphism. By analyzing the distribution of the polymorphic loci in all the examined individuals,we discovered a locus that was only existing among male individuals and was absent among females. In consistent with this observation, no male individual was detected in the artificial gynogenetic group of theE. ilishaeformis, and the locus was not detected in the genome of all examined gynogenetic individuals. The results suggested that sex determination and differentiation in this species may be strictly controlled by genes located in that specific region, and that gender difference locus we detected can be used as a male-specific marker. Our statistical analysis showed that the percentage of polymorphic loci of Taihu strain was 57.99%, the observed number of alleles of Taihu strain was 1.5799±0.4943, the effective number of alleles of Taihu strain was 1.3859±0.3971, and the Shannon’s information index of the population was 0.3221±0.2987. These data indicated that the genetic diversity of Taihu strain is moderate and is maintained at a similar level (P=51.21%,Na=1.512±0.500,Ne=1.252±0.371,I=0.218±0.275) compared with the results obtained 10 years ago.
Erythroculter ilishaeformis; Genome analysis; Sexual dimorphism; Genetic diversity
Q346+.5
A
1000-3207(2017)06-1200-07
2016-12-27;
2017-04-26
浙江省重点研发计划(2016C02055-1)资助 [Supported by Zhejiang Science and Technology Major Program (2016C02055-1)]
刘祥芳(1991—), 女, 山东日照人; 硕士; 主要从事鱼类遗传学研究。E-mail: 1559023141@qq.com
罗琛, 男, 教授, 博士生导师; 主要从事细胞与发育生物学研究。E-mail: luoc@zju.edu.cn
10.7541/2017.149
