不同浓度盐胁迫下盐穗木叶片结构的比较观察
2016-11-26王虹齐政张富春
王虹,齐政, 张富春
(新疆生物资源与基因工程重点实验室/新疆大学生命科学与技术学院,乌鲁木齐 830046)
不同浓度盐胁迫下盐穗木叶片结构的比较观察
王虹,齐政, 张富春
(新疆生物资源与基因工程重点实验室/新疆大学生命科学与技术学院,乌鲁木齐 830046)
【目的】盐穗木叶片退化成鳞片状,在植株的光合作用中不是起主要作用,但其对逆境的变化很敏感,研究盐胁迫下盐穗木叶片结构变化,找出其最佳盐浓度范围。【方法】可用不同盐浓度胁迫下,切片后显微镜观察盐穗木叶片细胞结构的变化。【结果】随着盐浓度的增加,盐穗木叶片逐渐变厚,细胞排列由疏松到紧密,含晶细胞和储水细胞大而多,表皮细胞壁向外呈乳突状凸起逐渐增多,可以充分的吸收水分,而叶片表皮细胞壁外凸又由乳突状到毛状,细胞中水分的减少。叶表皮和气孔中有较多晶体,盐穗木叶片上的气孔器与表皮细胞的角质层均可以向外泌盐。【结论】盐穗木是一种聚盐和泌盐混合形式的盐生植物。在盐浓度在300~500 mmol/L下,盐穗木叶片细胞结构较正常,是其生长的最佳盐浓度范围。
盐穗木;盐胁迫;叶片;结构;解剖学
0 引 言
【研究意义】盐穗木(Halostachyscaspica(M.B.) C. A. Mey.)是藜科(Chenopodiaceae)盐穗木属(HalostachysC.A.Mey.)的半灌木盐生植物[1],在新疆各地均有分布。盐穗木生长于荒漠地区盐碱地,盐湖边及河岸,绿期长,是具有重要生态价值的盐生植物,常与耐盐碱及耐干旱的灌木、草本植物形成群落及复合群落,也常成为盐生植物群落的建群种或优势种[2]。因此,对盐穗木叶片进行不同浓度的盐胁迫,观察分析其结构的变化,系统的研究盐穗木叶片的其他特性和功能,对恢复生态建设、改善土壤条件具有重要意义。【前人研究进展】对盐生、旱生植物形态结构的研究已有很多,角度也各不相同[3-8]。近年来,对盐穗木的研究主要集中在基因工程、分子生物学等领域[9-14];赵恩峰等[15]对盐穗木在盐胁迫下的水势、CAT、MDA、电导率、叶绿素含量等生理指标进行了分析,而这些研究都是针对盐穗木同化枝进行的,而对其叶片的结构基本上是一带而过[16,17]。【本研究切入点】虽然盐穗木叶片退化成鳞片状,在植株的光合作用中不是起主要作用,但在当年生的枝条上,盐穗木的叶片对生境的变化也是很敏感的。有关不同浓度盐胁迫下盐穗木叶片结构的研究未见文献报道。研究不同浓度盐胁迫下盐穗木叶片结构。【拟解决的关键问题】对不同浓度盐胁迫下(设置不同的盐梯度),盐穗木叶片生长7 d后,观察各组织结构和细胞的变化、盐晶体聚集的位置、盐晶体分泌的方式等进行观察比较,找出其盐穗木叶片生长的最佳盐浓度范围。
1 材料与方法
1.1 材 料
采集野外盐碱地(新疆乌鲁木齐市北五家渠103团)盐穗木种子。
1.2 方 法
种子带回实验室,播种于铺有基质(质量比为珍珠岩∶蛭石=3∶1,用双子叶植物营养液拌湿)的花盆中,在(24±2)℃,光照3 000 lx(350 μmol/(m2·s),16 h/8 h:昼/夜),相对湿度为40%~60%条件下,并施以1/4 MS(Murashige and Skoog )培养液培养。样本材料的盐梯度分别为0 mmol/L(空白对照)、100、300、500和700 mmol/L,胁迫时间为7 d。分别取经过这5个盐梯度处理的样品叶片,放入FAA固定液中固定,经酒精系列脱水,石蜡包埋后进行切片(厚7 μm)。切片经二甲苯脱蜡透明,番红—固绿对染,中性树胶封片,在显微镜下观察并摄影。
2 结果与分析
2.1 盐穗木叶片在0 mmol/L盐浓度下的结构
在0 mmol/L盐浓度下(图1:1~2),叶片细胞达到5~7层,叶表皮具有明显角质层,气孔器中有晶体;表皮细胞外凸,栅栏组织分化不明显,细胞排列疏松且不规则,叶绿体多分布在细胞壁上,颗粒不明显;薄壁细胞间隙大,不规则松散排列;维管束细小,不明显,数量较少,有少量细小的晶体在气孔器和薄壁细胞中;上表皮细胞下1~2层薄壁细胞不规则疏松排列。表1,图1
2.2 盐穗木叶片在100 mmol/L盐浓度下的结构
在100 mmol/L盐浓度下(图1:3~5),叶表皮细胞壁外凸泡状明显;角质层明显,薄壁细胞较规则镶嵌排列,细胞体积有所增大,栅栏组织细胞分化逐渐明显,形成较规则的大空腔,叶绿体逐渐增多,颗粒也增大,分布较均匀,维管束呈纵向条状排列,较小,数量少,晶体大多数聚集在靠近维管束的薄壁细胞中,上表皮细胞较大,排列较规则,有含晶细胞分布。图1
2.3 盐穗木叶片在300 mmol/L盐浓度下的结构
在300 mmol/L盐浓度下(图1:6~8),叶片细胞最厚达到8~10层,细胞呈纵向拉长;叶表皮细胞壁外凸呈泡状毛,内有盐积累,角质膜增厚,薄壁细胞体积变小,排列紧密,细胞内含晶体;栅栏组织排列较规则,细胞多为柱状,大空腔增多;叶绿体多,颗粒清晰,基本充满栅栏组织的第一层细胞中;维管束开始变粗,呈纵向条状排列;海绵组织不明显,细胞变小,数量也增多,晶体多分布在薄壁细胞壁和维管束中,上表皮细胞排列紧密,薄壁细胞变小,晶体分布多。表1,图1
2.4 盐穗木叶片在500 mmol/L盐浓度下的结构
在500 mmol/L盐浓度下(图2:9~11),叶片细胞最厚达到8~10层,叶表面细胞壁凸起呈毛状增多达到高峰,角质层浓厚;薄壁细胞排列不规则,含晶细胞、储水细胞体积大;栅栏组织呈柱状规则排列,较为紧密,空腔大而多,有簇晶积累,叶绿体浓密,颗粒大而清晰;分化不明显的海绵组织中细胞排列紧密;维管束呈纵向条状排列,数量较多,晶体几乎充满导管中;上表皮细胞排列不规则,储水细胞,薄壁细胞内有大量盐晶体大。表1,图2
2.5 盐穗木叶片在700 mmol/L盐浓度下的结构
在700 mmol/L盐浓度下(图2:12~14),叶片外表皮泡状毛的细胞壁破裂,角质层不均匀,薄壁细胞排列较紧密,细胞壁破裂,栅栏组织细胞排列较紧密但不规则,细胞壁破裂,空腔变小,叶绿体颗粒模糊,分布不均匀,维管束发达,数量多,晶体几乎充满所有导管;叶肉细胞中也都有晶体;上表皮细胞排列相对规则,薄壁细胞壁多数破裂。在有盐胁迫时,叶厚随盐浓度增大而减小,在此浓度下最小。表1,图2
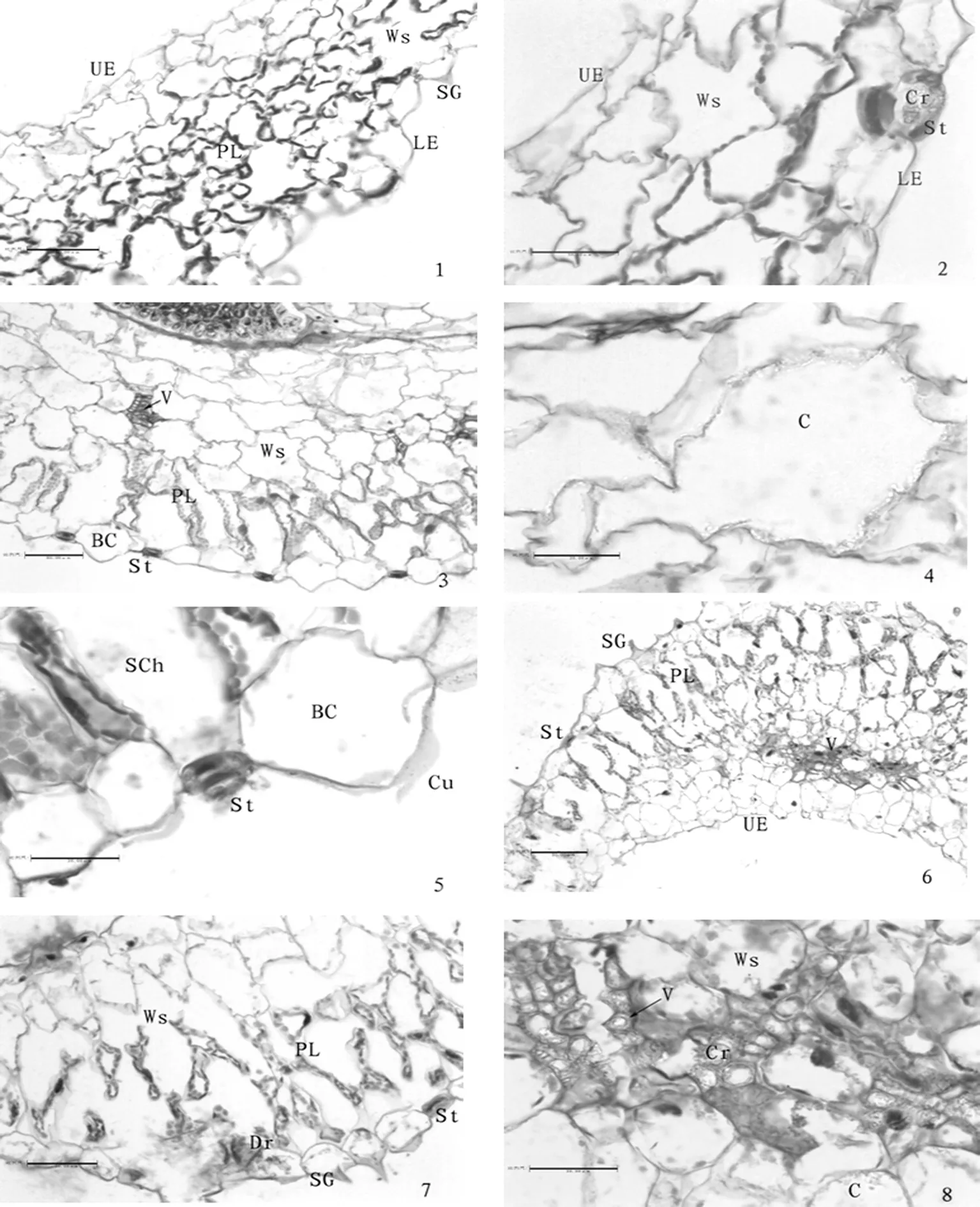
C.含晶细胞;Cu.角质膜;LE.下表皮;UE.上表皮;PL.栅栏组织;SG.盐腺;St.气孔器;V.维管束;Ws.储水组织;BC.泡状细胞;VH.泡状毛;Cr.晶体;Dr.簇晶;SC.沙晶;Ch.叶绿体;SCH.孔下室;PC.薄壁细胞
C.crystal cell;Cu.Cuticula;LE.lower epidermis;UE.upper epidermis ;PL.palisade tissue;SG. salt gland; St.Stoma;V.Vascular bundle;Ws.Water-storing tissue;BC.bulliform cells;VH.vesicular hairs;Cr.crystals;Dr.druses;SC.sand crystals;Ch.chloroplast; SCh.substomatic chamber.;PC.parenchymal cell
Ⅰ 1,2.盐穗木在0 mM(对照)下叶片的结构特征. 示气孔器中的晶体 ×200.×200. 3~5.盐穗木在100 mM盐胁迫下叶片的结构特征.示维管束、含晶细胞、泡状细胞及下陷的气孔器和孔下室.×100.×200.×400. 6~8.盐穗木在300 mM盐胁迫下叶片的结构特征. 示维管束、晶体、储水组织,叶的下表皮细胞有簇晶. ×100.×200. ×400.
PlateⅠ1,2.Leaf structure ofHalostachyscaspicaunder 0 mM salt concentration. Showing crystal in stoma. ×200.×200. 3-5.Leaf structure ofHalostachyscaspicaunder 100 mM salt concentration. Showing vascular bundle, crystal cells, bulliform cells, sunk stoma and cell under stoma. ×100.×200.×400. 6-8.Leaf structure ofHalostachyscaspicaunder 300 mM salt concentration. Showing vascular bundle, crystal and water-storing tissue,druses in under epidermal cells. ×100.×200.×400.
图1 盐穗木在0 mmol/L、100 mmol/L、300 mmol/L下叶片的结构特征
Fig.1 Leaf structure of Halostachys caspica under 0 mmol/L、100 mmol/L、300 mmol/L salt concentration
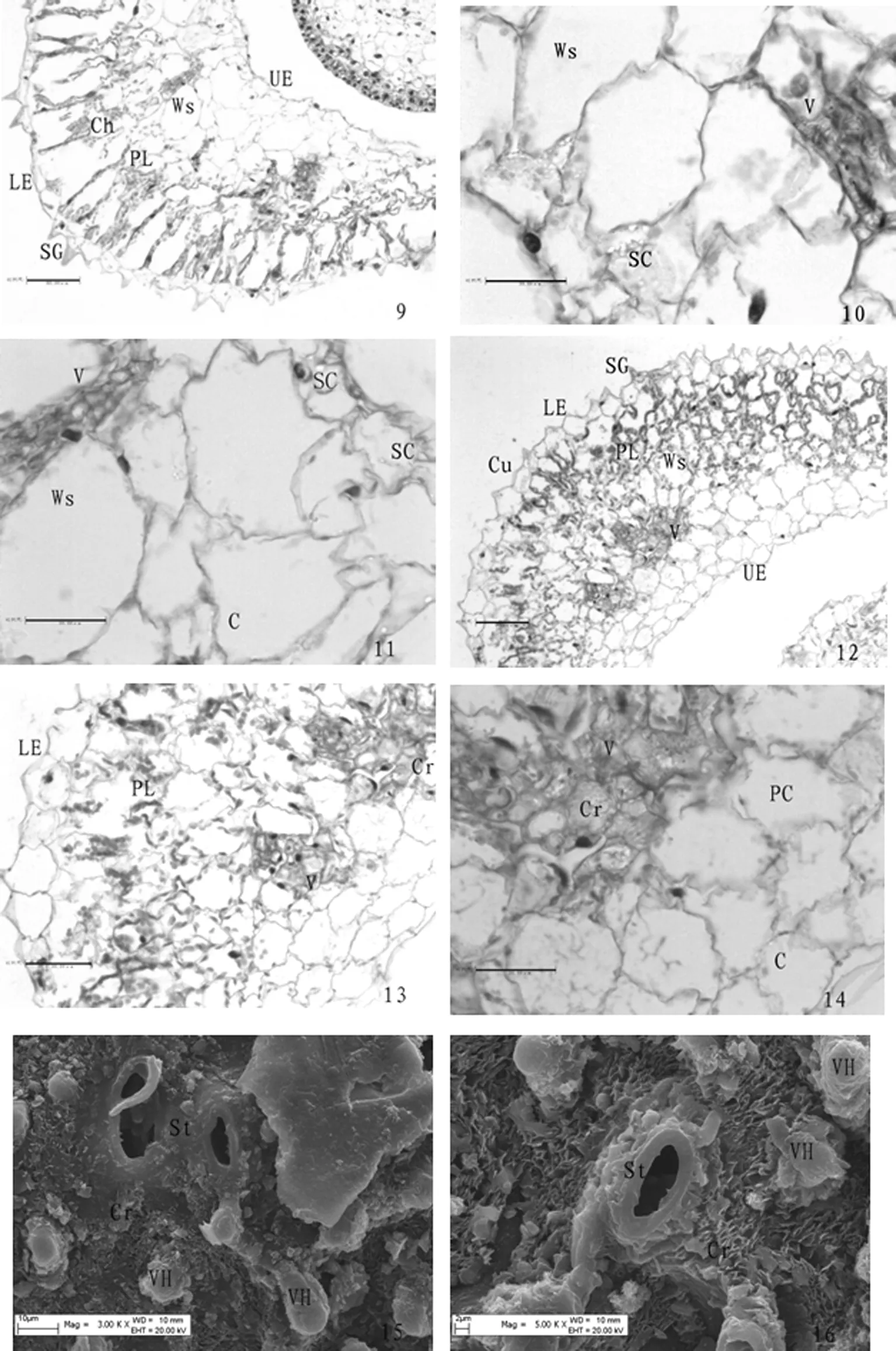
Ⅱ9~11.盐穗木在500 mM盐胁迫下叶片的结构特征. 示叶表皮细胞壁凸起呈毛状、栅栏组织、叶绿体、含晶细胞及沙晶. ×100.×200.×400. 12~14.盐穗木在700 mM盐胁迫下叶片的结构特征. 示泡状细胞壁、薄壁细胞壁的状态及晶体的分布.×100.×200.×400. 15,16.扫描电镜下,盐穗木叶的上、下表皮,示气孔器、盐结晶、泡状毛.×3000.×5000.
PlateⅡ9-11.Leaf structure ofHalostachyscaspicaunder 500 mM salt concentration. Showing palisade tissue, chloroplast, crystal cells, sand crystals and hair-like wall bump of epidermal cells. ×100.×200.×400. 12-14.Leaf structure ofHalostachyscaspicaunder 700 mM salt concentration. Showing the wall of bulliform cell, the wall of parenchyma cells and the distribution of crystal.×100.×200. ×400. 15,16. Upper and under epidermal ofHalostachyscaspicaunder SEM. Showing stoma, salt crystal and vesicular hairs. ×3000.×5000.
图2 盐穗木在500 mmol/L(对照)、700 mmol/L下叶片的结构特征
Fig.2 Leaf structure of Halostachys caspica under 500 mmol/L、700 mmol/L salt concentration
表1 盐穗木分别在不同盐浓度胁迫下叶片结构比较
Table 1 Comparative struture of Halostachys under different salt concentration stress

盐浓度(mmol/L)上表皮细胞厚度Thicknessofupperepidermiscell上表皮细胞宽度Widthofupperepidermiscell栅栏组织Palisadetissue海绵组织Spongytissue下表皮细胞厚度Thicknessoflowerepidermiscell下表皮细胞宽度Widthoflowerepidermiscell叶厚Thicknessofleaf图版Plate0(对照)(Contrast)6007~98725439~961112865~312647480~126092604~76583960~812930395~57796图1:1-21007555~189837057~1593331408~9815020392~364388817~140866040~1545664506~150044图1:3-53003682~87707287~1270025454~6839931671~560172457~43776861~1098762448~129061图1:6-85003906~108885248~1899931183~4779325897~529134367~80895592~914767797~112741图2:9-117005527~111195730~1115820223~4454614829~258251667~62516628~903641454~75131图2:12-14
3 讨 论
植物的耐盐和耐旱性是一个复杂的综合性状,是由多因素共同作用的结果[18]。盐渍长期作用于植物,诱导盐生植物形态结构的发育出现一系列变化来适应盐胁迫。肖雯[19]指出,不同的植物种类、器官和盐分的类型也都影响植物形态结构的变化,分化出具有明显生态适应性的结构,如植物叶片表皮细胞壁外凸形成泡状细胞、表面有角质膜、气孔器下陷并有明显的孔下室、具有含晶细胞、泌盐结构等等[17,20]。赵曼容等[21](1993)认为,典型盐地植物的形态及结构通常都可看作是对盐度的适应性。
3.1 盐穗木的最适盐浓度
以往学者对盐穗木细胞结构的研究,是将叶片与同化枝上放在一起观察分析,可能是因为盐穗木叶片很小,在植物的光合作用中作用不大[16,17]。但是作为植物中重要的营养器官之一,盐穗木叶片在不同盐浓度胁迫下,其细胞结构也发生了明显的变化,说明其对生境的变化也有同样的敏感性。结果表明:(1)虽然盐浓度为0 mmol/L盐浓度时没有盐胁迫,盐穗木叶片细胞中仍然有晶体存在长,这可能是生长的土壤和水中也含有少量盐分子的缘故。在0,100 mmol/L盐浓度下,盐穗木叶片较薄,生长较弱,说明在盐浓度很低时并不利于盐穗木叶片的生长发育。(2)300~500 mmol/L盐浓度下,盐穗木叶片这时呈现出典型盐生、旱生植物叶片的特点[17,20],而且生长发育很好。赵恩峰等[15]也指出这时的盐穗木植株超氧阴离子(O)含量、SOD活性及电导率呈现最低值;丙二醛(MDA)含量也较低,而叶绿素含量则较高,盐穗木植株在该浓度范围内生长良好,生物量高。(3)盐穗木在700 mmol/L盐浓度胁迫下,叶片中栅栏组织逐渐由柱状细胞变为小圆形,叶绿体也逐渐由浓密变稀疏,颗粒变的模糊,分布也不均匀,并且薄壁细胞变大,细胞壁也变得模糊甚至破裂,Qi C H等[22]认为这是因为过量的盐离子对植物造成的毒害,甚至导致植物细胞死亡。因此在不同盐浓度胁迫下,盐穗木叶片细胞结构的变化,从另外一个角度验证了其生理特性变化的结论[15]。300与500 mmol/L的盐浓度是盐穗木生长的最适盐浓度,盐浓度太小或太大都会影响盐穗木叶片结构和细胞的正常发育生长。
3.2 与环境的适应性
通过对盐穗木叶片结构切片的观察发现,随着盐浓度逐渐增加,在100 mmol/L时叶片厚度达到最大(表1),之后叶片厚度及细胞体积又逐渐减小,这与李瑞梅等[4]对海马齿的研究结论基本一致。较高浓度盐胁迫下,叶片表皮角质膜增厚,这与B.P.Strogonov[23]和Y. Waisel[24]所得出来的结论一致。在100~700 mmol/L盐浓度时,盐结晶主要都是分布在盐穗木叶的维管束、海绵组织及上表皮的薄壁细胞中,多为小颗粒状的沙晶,而在栅栏组织及下表皮细胞中的盐结晶较少,多为大的簇晶,随着盐浓度的增加,可观察出植物体内的过多的盐分累积在泡状毛的液泡中,盐穗木叶片在不同盐浓度胁迫下,随着盐浓度和胁迫时间的增加,叶细胞层数逐渐增加,表皮细胞壁向外呈乳突状凸起,而且数目也越多,凸起的状态也越强烈,甚至整个表皮细胞壁向外凸起,以增大细胞的表面积和表皮细胞的体积,充分的吸收水分,来抵御干旱的环境。然而,叶片表皮细胞壁外凸又由乳突状到毛状,则是水分丧失的征兆,而不是减少水分蒸腾作用的结构[25],说明此时的细胞中水分正在减少,表明这是叶片适应盐生、旱生环境的结果[26]。费贯清[27]认为植物存在的三种抗盐机理:避让、忍耐和缓解,是盐生植物和盐胁迫下植物发生较为典型的表层结构的变化,而气孔器下发达的孔下室,也是为了防止水分的蒸发。
3.3 盐穗木的盐生植物类型
盐穗木叶片内的大量含晶细胞是植物对盐碱的一种积累形式[6,17],随着盐浓度的增加,盐穗木叶下表皮的细胞表面的盐结晶及泡状毛均比上表皮的多,泡状毛外也有很厚的盐结晶层(图2:15,16),说明盐通过角质膜上的小孔被排出盐穗木的叶片;而叶片上的气孔器凸起呈张开状,最后失去了闭合的功能,气孔内也有盐结晶,可能气孔器与表皮细胞的角质膜一样均可以向外泌盐,但在植物体内,特别是在叶片的薄壁细胞和维管束中有大量盐的积累,叶片通过储水细胞稀释盐浓度的方式使吸收到体内的盐分不致发生毒害,从而体现出较高的耐盐性[11],说明盐穗木既有泌盐功能又有聚盐功能,因此,邓彦斌等[17]将盐穗木归类为泌盐和聚盐植物的一种形式。盐穗木叶片所具备的这些盐生、旱生植物结构特性,可起到良好的保护作用,使其安全的完成泌盐过程,能在较高盐浓度的土壤环境下生长,而避免遭受严酷环境的损伤,能最大限度的抵御恶劣环境造成的伤害而生存下来。
4 结 论
随着盐浓度的增加,盐穗木叶片逐渐变厚,细胞排列由疏松到紧密,含晶细胞和储水细胞大而多;表皮细胞壁向外呈乳突状凸起逐渐增多,可以充分的吸收水分,而叶片表皮细胞壁外凸又由乳突状到毛状,说明细胞中水分减少了,盐穗木叶的细胞结构在水分条件好时可以尽可能的储存水分,而在干旱时又可以转换成保水模式,以便抵御干旱环境的影响。另外,盐穗木叶表皮和气孔中有较多晶体,表明盐穗木叶片上的气孔器与表皮细胞的角质层均可以向外泌盐;说明盐穗木是同时具备泌盐和聚盐形式的盐生植物;其生长的最佳盐浓度范围在300~500 mmol/L,为全面系统的研究盐穗木的其他特性和功能提供一些科学数据,也对新疆广袤的干旱盐碱地区的土壤改善和生态恢复建设具有重要意义。
References)
[1] 毛祖美,刘建国,潘晓玲,等.新疆植物志(第二卷第一分册)[M]. 新疆科学卫生出版社,1994:13-14.
MAO Zu-mei, LIU Jian-guo, PAN Xiao-ling,et al. (1994).FloraofXinjiang. [M]. Urumqi: Xinjiang Science & Technology & Hygiene Publishing House: 13-14.(in Chinese)
[2] 涂锦娜,王绍明.盐穗木生态场土壤水盐及养分的空间异质性研究[D].石河子:石河子大学,2011.
Tu Jin-na, Wang Shao-ming.Studiesonspatialheterogeneityofecologicalfieldsoilmoisture,salinityandnutirentsofHalostachyscaspica. [D].Shihezi,Shihezi University,2011. (in Chinese)
[3] 赵可夫,李法曾,樊守金,等.中国的盐生植物[J]. 植物学通报,1999,16 (3):201-207.
ZHAO Ke-fu, LI Fa-zeng, FAN Shou-jin, et al. (1999). Halophytes In China [J].ChineseBulletinofBotany, 16 (3):201-207. (in Chinese)
[4] 李瑞梅,周广奇,符少萍,等.盐胁迫下海马齿叶片结构变化[J]. 西北植物学报,2010,30(2):287-292.
LI Rui-mei, ZHOU Guang-qi, FU Shao-ping, et al. (2010).Leaf Anatomical Structure ofSesuviumportulacastrumL. under Salt Stress [J].ActaBotanicaBoreali-OccidentaliaSinica,30(2):287-292. (in Chinese)
[5] 黄振英,吴鸿,胡正海.新疆10种沙生植物旱生结构的解剖学研究[J].西北植物学报,1995,15(6):56-61.
HUANG Zhen-ying, WU Hong, HU Zheng-hai. (1995).An Anatomical Study On Xeromorphic Structures of the Psammophytes in Xinjiang [J].ActaBotanicaBoreali-OccidentaliaSinica, 15(6):56-61. (in Chinese)
[6] 王虹,邓彦斌,徐秀珍,等.新疆10种旱生、盐生植物的解剖学研究[J].新疆大学学报(自然科学版),1998,15(4):67-73.
WANG hong, DENG Yan-bin, XU Xiu-zhen, et al. (1998). Anatomical Studies On Ten Xerophytes and Halophytes in Xinjiang [J].JournalofXinjiangUniversity(NaturalScienceEdition),15(4):67-73. (in Chinese)
[7] 周桂玲,迪利夏提,安争夕,等.新疆滨藜属植物叶表皮微形态学及叶的比较解剖学研究[J].干旱区研究, 1995,12(3):34-37.
ZHOU Gui-ling, Dilixiati, AN Zheng-xi, et al. (1995). Study on the Micromorphology of Leaf Epidermis and the Comparative Anatomy of Leaf of Atriplex in Xinjiang [J].AridZoneResearch, 12(3):34-37. (in Chinese)
[8] 陈玲,兰海燕.北疆荒漠几种盐生(耐盐)植物抗逆附属结构的初步研究[J]. 广西植物,2012,32(5):686-693.
CHEN Ling, LAN Hai-yan. (2010). Four halophytes (or salt-tolerant species) and their accessory structures of leaf related to adversity tolerance in the desert of northern Xinjiang [J].Guihaia, 32(5):686-693. (in Chinese)
[9] 彭丹,张霞,张富春,等.盐穗木过氧化氢酶基因的克隆与功能分析[J]. 西北植物报,2013,33(10):1 933-1 939.
PENG Dan, ZHANG Xia, ZHANG Fu-chun, et al. (2013). Cloning and Functional Identification of Catalase Gene inHalostachyscaspica[J].ActaBotanicaBoreali-OccidentaliaSinica, 33(10):1,933-1,939. (in Chinese)
[10] 张霞,张富春.盐生植物盐穗木悬浮细胞系的建立[J]. 新疆农业科学,2013,50(9):1 718-1 723.
ZHANG Xia, ZHANG Fu-chun. (2013). Establishment of Suspension Cell Line ofHalostachyscaspica[J].XinjiangAgriculturalSciences, 50(9):1,718-1,723. (in Chinese)
[11] 郝晓燕,张毓露,足木热木,等.盐穗木HcPS-H基因真核表达载体的构建及转化[J]. 新疆农业科学,2012,49(12):2 263-2 268.
HAO Xiao-yan, ZHANG Liu-lu, Zumuremu, et al. (2012). Construction and Transformation of Eukaryotic Express Vector of HcPS H Gene fromHalostachyscaspica[J].XinjiangAgriculturalSciences, 49(12):2,263-2,268. (in Chinese)
[12] 木合热皮亚·艾尔肯,张富春.盐穗木HcVP1转基因棉花耐盐性的生理生化分析[J]. 新疆农业科学,2013,50(6):1 016-1 020.
Muherepiya Aierken,ZHANG Fu-chun. (2013).Physiological and Biochemical Analysis for the Salt Tolerance of Transgenic Cotton Transformed by the Vacuolar H+ 一Pyrophosphatase Gene(VP1) fromHalostachyscaspica[J].XinjiangAgriculturalSciences, 50(6):1,016-1,020. (in Chinese)
[13] 王志才,廖茂森,木合热皮业·艾尔肯,等. 盐穗木耐盐基因通过花粉管通道法对棉花遗传转化的研究[J]. 分子植物育种,2011,9(2):180-184.
WANG Zhi-cai, LIAO Mao-sen, Muherepiya Aierken, et al. (2011).Study on Pollen-tube Pathway Mediated Cotton (Gossypiumhirsutum) Transformation with Salt-tolerant Gene fromHalostachyscaspica[J].MolecularPlantBreeding, 9(2):180-184. (in Chinese)
[14] 姚世响,陈莎莎,徐栋生,等.不同盐胁迫对新疆耐盐植物藜叶片钾、钠元素含量及相关基因表达的影响[J]. 光谱学与光谱分析,2010,30(8):2 281-2 284.
YAO Shi-xiang, CHEN Sha-sha, XU Dong-sheng, et al. (2010). Effects of Different Salinity Stress on K, Na Content and Relevant Genes Expression in Leaves ofChenopodiumAlbumin Xinjiang [J].SpectroscopyandSpectralAnalysis,30(8):2,281-2,284. (in Chinese)
[15] 赵恩峰,李玲,曾幼玲.盐胁迫对盐生植物盐穗木生理特性的变化分析[J]. 新疆农业科学,2012,49(4):716-722.
ZHAO En-feng, LI Ling, ZENG You-ling. (2012).Physiological Responses of theHalophyteHalostachyscaspica to Salt Stress[J].XinjiangAgriculturalSciences,49(4):716-722. (in Chinese)
[16] 周玲玲,冯元忠,刘萍,等.新疆六种盐生植物的解剖学研究[J].石河子大学学报(自然科学版),2002,6(3):217-221.
ZHOU Ling-ling, FEN Yuan-zhong, LIU Ping, et al. (2002).Anatomical Studies on Six Halophytes in Xinjiang [J].JournalofShiheziUniversity(NaturalScience),6(3):217-221. (in Chinese)
[17] 邓彦斌,姜彦成,刘健,等.新疆10种藜科植物叶片和同化枝的旱生和盐生结构的研究[J]. 植物生态学报,1998, 22(2): 164-170.
DENG Yan-bin, JIANG Yan-cheng, LIU Jian, et al. (1998). The Xeromorphic and Saline Morphic Structure of Leaves and Assimilating branches in ten chenopodiacea species in Xinjiang [J].ActaPhytoecologicaSinica, 22(2): 164-170. (in Chinese)
[18] 刘滨,郑丽萍,任树勇,等.宁夏10种观赏灌木叶片解剖结构及其抗旱性综合评价[J]. 西北植物学报, 2013,33(9):1 808-1 816.
LIU Bin, ZHENG Li-ping, REN Shu-yong, et al. (2013).Drought Resistance Study of 10 Major Ornamental Shrub in Ningxia [J],ActaBot.Boreal.-Occident.Sinica, 33(9):1,808-1,816. (in Chinese)
[19] 肖雯.几种盐生植物抗盐生态适应性的研究[D].兰州,甘肃农业大学,1998:1-8.
Xiao Wen.Studiesonecologicaladaptationcharacteristicofseveralhalophytes[D].Lanzhou,Gansu Agricultural University,1998:1-8. (in Chinese)
[20] 肖雯.五种盐生植物营养器官显微结构观察研究 [J].甘肃农业大学学报,2002,37(4):421-427.
XIAO Wen. (2002). Microstructures in the nutritional organs of five halophytes [J].JournalofGansuAgriculturalUniversity,37(4):421-427.
[21] 赵曼容,贾恢先.几种典型盐地植物超微结构的研究[J].干旱区资源与环境,1993,3(3):64-68.
ZHAO Man-rong, JIA Hui-xian. (1993). Studies on Ultrastructure of several typical halophytes [J].JournalofAridLandResourcesandEnvironment,3(3):64-68. (in Chinese)
[22] Qi, C. H., Chen, M., Song, J., & Wang, B. S. (2009). Increase in aquaporin activity is involved in leaf succulence of the euhalophyte suaeda salsa, under salinity.PlantScience, 176(2), 200-205.
[23] Garg, B. K., Gupta, I. C., Joshi, D. C., & Kumar, D. (2000).Physiologicalbasisofsalttoleranceinplants.
[24] Waisel, Y. (1972).BiologyofHalophytes.New York: Academic Press.
[25] Shields, L. M. (1950). Leaf xeromorphy as related to physiological and structural influences.BotanicalReview, 16(8):399-447.
[26] AoFahn,植物解剖学[M]. 吴树明,等译. 天津:南开大学出版社,1990.
WU Shu-ming, et al. (1990).PlantAnatomyby A. Fahn [M],Tianjing:Nankai University Press. (in Chinese)
[27] 费贯清.盐生植物抗盐适应特征的研究[D].兰州:兰州大学,2005.
FEI Guan-qing. (2005).Studiesonsaltresistanceadaptabilityofhalophytes[D]. Lanzhou University,Lanzhou. (in Chinese)
Fund project:Supported by the earmark funds for national "973" plan preliminary research projects (2012CB722204); special funds for Xinjiang key laboratories (2014KL001)and open projects for Xinjiang key laboratories(XJDX0201-2014-04)
Leaf Anatomical Structure of Halostachys caspica under Different Concentrations of Salt Stress
WANG Hong, QI Zheng, ZHANG Fu-chun
(Xinjiang Key Laboratory of Biological Resources and Genetic Engineering, College of Life ScienceandTechnology,XinjiangUniversity,Urumqi830046,China)
【Objective】 Although leaves ofHalostachyscaspicadegenerate into a scaly shape and don't play a major role in plant photosynthesis, but they are very sensitive to the changes of environment.【Method】As an important organ, the leaf cell structure was observed under different salt concentration stresses by microscopes. 【Result】With the increase of salt concentration, the leaves ofH.caspicawere getting thicker progressively, cells arrangement from loose to tight, and crystal and storage cells became larger and more. Papillary-like bulge of epidermal cell wall showed a gradual increase in the outward convex shape, which could fully absorb the water. The variation in papillary-like to hair-like bulge of leaf epidermal cell wall indicated that water content in cells was decreased. More crystal in the leaf epidermal and stomata showed that the cuticle of epidermal cells and stomata ofH.caspicacould secrete salt.【Conclusion】H.caspicais an excessive mixed halophyte plant both aggregating salt and salt secreting. 300 mmol/L-500 mmol/L salt concentrations are the best salt concentrations for the growing ofH.caspica, and leaf cell structure is normal under those salt concentrations.
Halostachyscaspica; salt stress; leaf; structures; anatomy
10.6048/j.issn.1001-4330.2016.11.018
2016-06-13
国家“973”计划前期研究专项(2012CB722204);新疆维吾尔自治区重点实验室专项资金(2014KL001);新疆维吾尔自治区重点实验室开放项目(XJDX0201-2014-04)
王虹(1962-),女,重庆人,高级实验师,博士研究生,研究方向为植物形态解剖、植物分类、植物细胞,(E-mail)wanghong211@163.com
张富春(1962-),男,河南人,博士,博士生导师,研究方向为分子生物学,(E-mail)zfcxju@xju.edu.cn
Q944
A
1001-4330(2016)11-2098-08
