不同砧木对黄果柑果实有机酸含量和酸代谢相关酶活性及基因表达的影响
2016-09-23曹淑燕古咸杰李清南汪志辉
曹淑燕,荣 毅,古咸杰,李清南,廖 玲,叶 霜,邱 霞,汪志辉,2
(1.四川农业大学 园艺学院,四川 成都 611130;2.四川农业大学 果蔬研究所,四川 成都 611130)
不同砧木对黄果柑果实有机酸含量和酸代谢相关酶活性及基因表达的影响
曹淑燕1,荣毅1,古咸杰1,李清南1,廖玲1,叶霜1,邱霞1,汪志辉1,2
(1.四川农业大学 园艺学院,四川 成都611130;2.四川农业大学 果蔬研究所,四川 成都611130)
为了探究不同砧木对黄果柑果实有机酸含量、酸代谢相关酶活性及基因表达的影响,为黄果柑砧木的选择提供理论依据。以枳壳、红橘、香橙为砧木,黄果柑实生苗为对照进行研究。黄果柑果实以积累柠檬酸为主,香橙砧能够最有效地降低果实有机酸含量,成熟期比实生苗低24.35%;嫁接能够一定程度的降低CS活性,而对PEPC、MDH活性的影响较小;花后250~330 d,ACO和NADP-IDH活性均表现出香橙砧>枳壳砧>红橘砧>CK的趋势;除实生苗外,黄果柑NADP-IDH表达水平与总酸含量间呈显著性负相关,而CS、MDH、ACO与总酸含量均未表现出显著相关性。香橙是黄果柑理想的砧木品种,砧木间果实有机酸差异是ACO和NADP-IDH活性共同作用的结果,且NADP-IDH基因的表达可能是影响不同砧木黄果柑果实中有机酸含量的关键限制因子。
黄果柑;砧木;有机酸;酶活性;基因表达
黄果柑(Citruscultivarcv.Huangguogan)为芸香科(Rutacese)柑橘亚科(Subfamily aurantioideae)植物,是我国具有自主产权的优良杂交柑橘[1]。其果实翌年3-5月成熟,具有极晚熟、无核、优质、丰产等优良经济性状[2]。良好的砧穗组合有2种。近年来,石棉县引进香橙作为新的黄果柑砧木,但3种砧木对黄果柑果实品质的影响尚不明确。
柑橘果实中所含的糖分、有机酸含量及其比值是决定柑橘果实风味的重要指标[3-4],决定了柑橘的营养和商业价值[5]。多数柑橘品种在成熟期糖酸比较低,严重影响了柑橘产业的发展[6]。相对来说,果实内糖含量的变化幅度较小,而有机酸的变化幅度较大。因此,对决定糖酸比大小而言,有机酸的影响更为重要[7]。柠檬酸合成酶(CS)、磷酸烯醇式丙酮酸羧化酶(PEPC)、顺-乌头酸酶(ACO)、异柠檬酸脱氢酶(IDH)和苹果酸脱氢酶(MDH)[8]等作为有机酸代谢的关键酶,通过三羧酸循环途径参与果实有机酸的代谢。砧木对柑橘的生长量[9]、果实品质[10-11]、抗逆性[12-13]等具有很大的影响作用。至今,对于柑橘果实有机酸代谢的研究主要集中在成熟过程中柠檬酸的代谢机理[14]、品种间的代谢差异[15]和胁迫处理等对代谢酶活性及基因表达的影响[16]上,有关砧木影响酸代谢的研究较少。通过2012-2014年的观察记录发现,不同砧木黄果柑的可滴定酸含量差异显著,但是上述3种砧木对黄果柑有机酸含量、相关酶活性及基因表达方面的研究尚未见报道。
本试验研究了不同砧木对黄果柑有机酸含量、酸代谢相关酶活性及基因表达的影响,从而了解其影响机理,最终为筛选出黄果柑的优良砧木,实现黄果柑产业的提质增效提供理论基础。
1 材料和方法
1.1试验材料
1.1.1试验地条件试验地位于四川省石棉县黄果柑栽培标准示范园,年均气温17 ℃,年均积温5 468 ℃,年均无霜期326 d,年日照数1 242.9 h,年均降雨量778.3 mm,属中亚热带干热河谷气候类型,壤土。
1.1.2试验材料以枳壳[Poncirustrifoliate(L.)Raf]、红橘(CitrusreticulataBlanco)和香橙[C.junos(sieb.) Tanaka]3种砧木嫁接的黄果柑为试材,黄果柑实生苗为对照。每种砧木选择树势、树载、管理方式基本一致的5年生健壮黄果柑3株,进行果实的采样与相关指标的测定。
1.2试验方法1.2.1采样方法2014年11月15日(花后210 d,果实转色期)至2015年3月15日,每隔20 d左右,在树冠的外围东、南、西、北4个方位,随机摘取生长正常、大小均匀、无病虫害的黄果柑果实各2个,每株树采8个,用液氮立即带回实验室,进行测定。
1.2.2黄果柑果实有机酸含量的测定参照郭燕[17]和肖玉明[13]的方法进行果实有机酸的提取与测定(高效液相色谱法)。
1.2.3酸代谢相关酶活性的测定参照 Hirai等[18]的方法进行酶液的制备,并用Srene法测定酶活性。
1.2.4总RNA的提取及cDNA第一链的合成RNA提取与纯化方法参照刘庆[19]和Zhang等[20]的方法略有改进。提取的黄果柑果实RNA用琼脂糖凝胶电泳和紫外分光光度计分别进行质量检测和浓度分析。用DNase去除残留的DNA杂质,然后严格地按照PrimeSciptTMRT reagent Kit (DRR037A) 反转录试剂盒(购自宝生物工程大连有限公司)说明书进行反转录,合成后续试验所需的cDNA。3次生物学重复。
1.2.5实时荧光定量PCR分析实时荧光定量PCR引物设计:根据笔者克隆的黄果柑相关基因序列片段(登录号:KU319552、KU319556、KU319555、KU319553、KU319557、GQ389668.1),运用Primer 5.0软件进行引物设计,并进行扩增筛选,结果见表1。
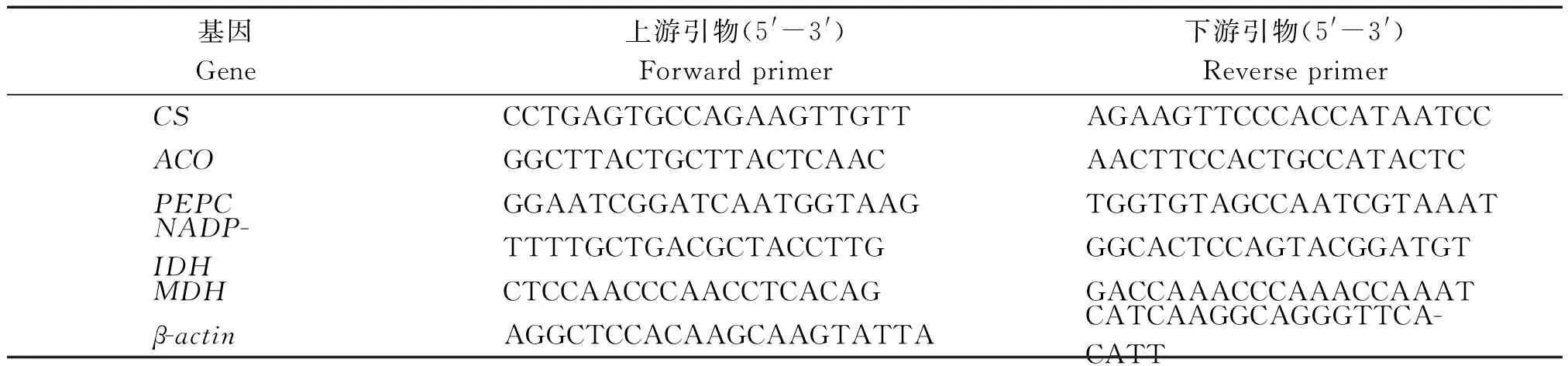
表1 黄果柑实时荧光定量PCR引物序列
基因的定量表达分析:以β-actin基因为内参基因进行实时荧光定量PCR分析,每个样品设3次生物学重复。反应根据SYBR®Rremix Ex TaqTM(Perfect Real Time)荧光定量试剂盒(购自宝生物工程大连有限公司)进行。反应体系:cDNA(1.5 μg) 2 μL;2×SYBR Green PCR Master Mix 12.5 μL;正、反向引物(10 μmol/L)各1 μL;ddH2O 8.5 μL,总体系为25 μL。反应程序:95 ℃,5 min;95 ℃,10 s;58 ℃,15 s;72 ℃,20 s(第2至4步进行40个循环)。溶解曲线在PCR运行完毕后立即开始。
1.2.6统计分析试验数据用Excel、SPSS 16.0软件进行处理分析。
2 结果与分析
2.1砧木对黄果柑有机酸含量的影响
由图1看出:不同砧木黄果柑柠檬酸含量与总酸含量变化趋势一致,呈先上升后下降的趋势,且柠檬酸含量占总酸含量的90.45%~96.79%,说明黄果柑果实以积累柠檬酸为主;3种砧木与实生苗相比,酸含量均有所下降,成熟期总酸含量分别降低13.17%,6.93%和24.35%,说明香橙砧木能够最有效地降低酸含量;CK、枳壳砧、红橘砧有机酸含量在花后230 d达到最高值,之后逐渐分解直到成熟,香橙砧最高值出现在花后250 d,说明香橙砧木能够推迟黄果柑的成熟;苹果酸和奎宁酸含量很低,整体呈现下降趋势,不同砧木之间无明显的规律性。
2.2砧木对黄果柑有机酸代谢相关酶活性的影响
2.2.1砧木对黄果柑有机酸合成酶类活性的影响由图2可知,不同砧木CS活性(以鲜质量计)在花后250 d达到峰值,之后缓慢下降,但仍维持在一个较高的活性水平,说明黄果柑有机酸在花后250 d后的快速降解受CS活性下降的影响较小;除了花后270 d,CS活性均表现为CK最高,但是3种砧木之间CS活性在各个时期表现出不同的差异情况,无明显规律,说明嫁接能够降低黄果柑CS活性,但3种砧木间有机酸含量的差异不是由CS活性的差异造成;不同砧木PEPC活性(以鲜质量计)在花后210~330 d均呈现快速下降的趋势,但各时期,处理之间的PEPC活性差异情况无明显规律,说明黄果柑成熟过程中PEPC酶活性的下降,导致果实有机酸含量下降,但不同砧木之间有机酸含量的差异形成与PEPC活性无关;MDH活性在花后250 d达到峰值,之后缓慢降低,花后330 d活性稍有提高,但处理之间MDH活性(以鲜质量计)在不同时期差异情况无明显变化规律,说明MDH活性的降低使得黄果柑有机酸含量降低,但不同砧木之间有机酸含量的差异与MDH活性关系不大。

图1 不同砧木对黄果柑柠檬酸、苹果酸、奎宁酸、总酸含量的影响
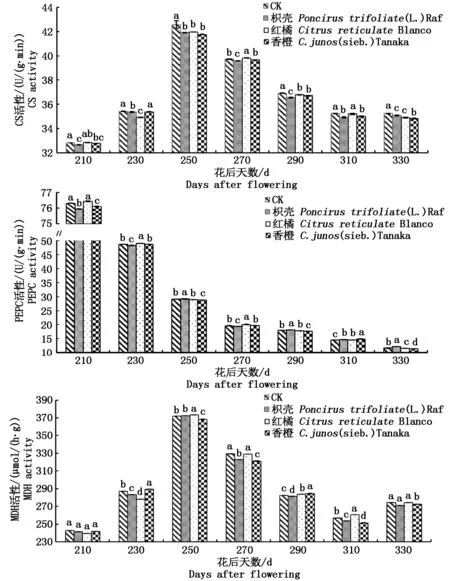
同一时期进行LSD检验,小写字母代表0.05差异水平。图3~5同。
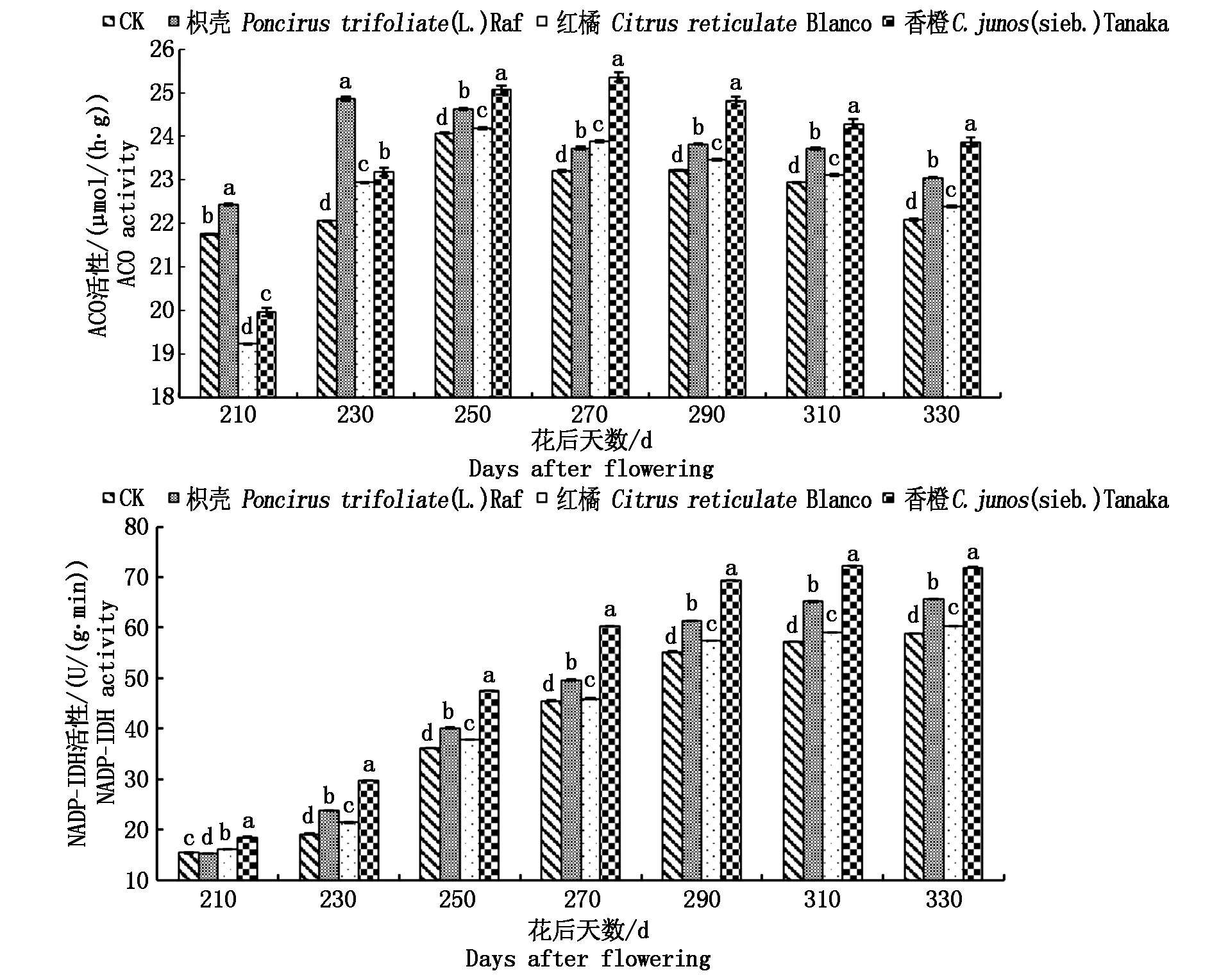
图3 不同砧木对黄果柑ACO和NADP-IDH活性的影响
2.2.2砧木对黄果柑分解酶类活性的影响由图3可以看出:不同砧木黄果柑ACO活性(以鲜质量计)呈现先上升后稍微下降的趋势,相同时期,各处理差异显著,且花后250~330 d均表现为香橙砧>枳壳砧>红橘砧>CK,说明嫁接能够提高ACO活性,从而降低有机酸含量,其中香橙砧木能够更好地提高ACO活性;NADP-IDH活性(以鲜质量计)随着黄果柑果实的成熟,呈现上升的趋势,同一时期差异显著,且花后230~330 d均表现为香橙砧>枳壳砧>红橘砧>CK,说明嫁接同样能够提高NADP-IDH的活性,且提高能力依次为香橙砧>枳壳砧>红橘砧>CK。综合说明,不同砧木黄果柑果实有机酸含量的差异,是ACO和NADP-IDH活性共同作用的结果。
2.3砧木对黄果柑有机酸代谢相关酶基因表达的影响
2.3.1砧木对黄果柑有机酸合成酶类基因表达的影响由图4可知:不同砧木合成酶类基因表达的变化趋势与酶活性的变化趋势一致,说明基因表达量的多少直接决定相应酶的活性强度。实生苗(CK)的CS基因表达量总体上高于其他砧木嫁接苗,说明嫁接能够在一定程度上抑制CS基因的表达;除个别时期,各个处理间PEPC和MDH相对表达量都无显著的差异,说明砧木基本不影响黄果柑果实PEPC、MDH的基因表达。

将实生苗(CK)花后210 d的数据作为1个表达水平计算各酶表达相对丰度。图5同。
2.3.2不同砧木对黄果柑有机酸分解酶类基因表达的影响由图5可以看出,在黄果柑成熟过程中,有机酸分解酶类基因的表达量,总体呈上升的趋势,说明在黄果柑成熟期有机酸的降解,取决于成熟期高的分解酶类基因的表达。枳壳砧果实的ACO表达峰值出现在花后230 d,CK和红橘砧的表达峰值出现在花后250 d,而香橙砧却在花后270 d,说明ACO基因的启动枳壳砧>CK和红橘砧>香橙砧,在一定程度上,枳壳砧木提早了黄果柑有机酸的分解,香橙砧木延迟了有机酸的分解;除了花后210 d,砧木间NADP-IDH的表达量差异显著,且香橙砧>枳壳砧>红橘砧>CK,说明嫁接能够改变黄果柑果实NADP-IDH的表达,导致果实相应酶活性发生变化,最终导致有机酸含量出现差异。
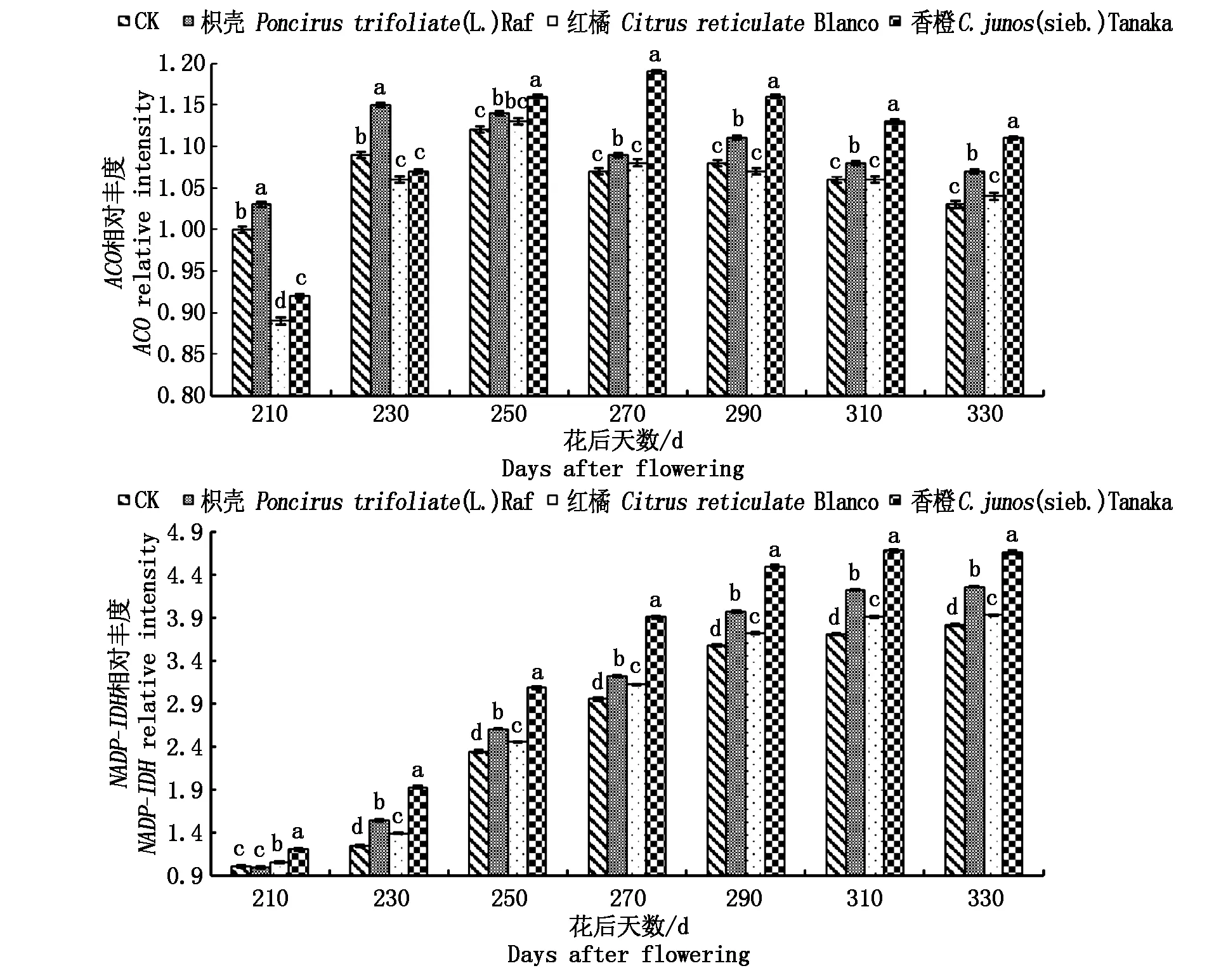
图5 不同砧木对顺乌头酸酶、异柠檬酸脱氢基因表达的影响
2.3.3不同砧木黄果柑有机酸代谢相关酶基因表达与总酸含量相关性分析将各处理有机酸代谢相关基因表达水平与总酸含量进行相关性分析,结果发现枳壳砧的果实中PEPC表达水平与总酸含量间呈显著性正相关,但在其他处理中相关性不显著;各处理的NADP-IDH表达水平与总酸含量间呈显著性负相关,且枳壳砧呈极显著负相关,而CS、MDH、ACO与总酸含量均未表现出显著相关性。上述结果说明NADP-IDH基因的表达是影响不同砧木黄果柑果实中有机酸含量的关键限制因子(表2)。
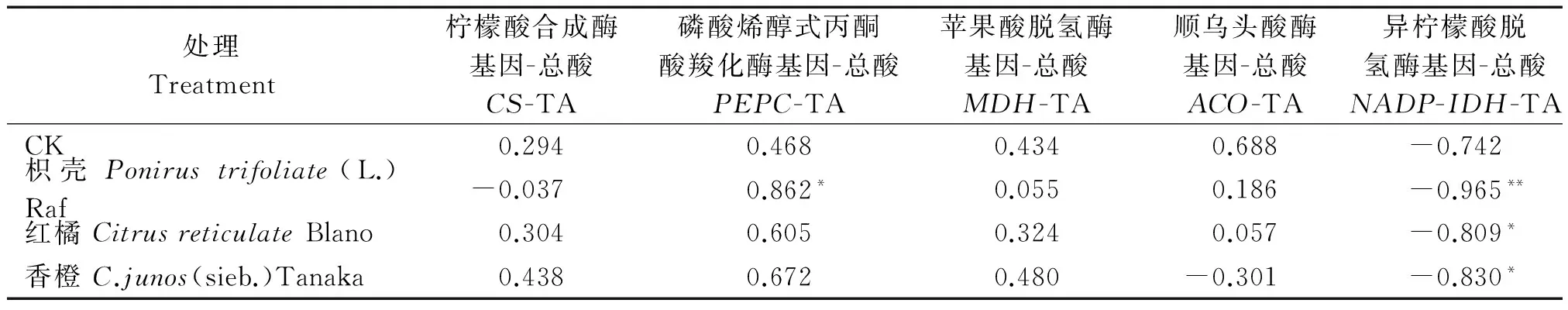
表2 不同砧木黄果柑有机酸代谢相关酶基因表达与总酸含量的相关系数
注:*和**分别表示在P<0.05 和P<0.01 水平下显著相关。
Note:*and**indicate significant difference atP<0.05 andP<0.01,respectively.
3 讨论
Yamaki[21]试验发现绝大多数柑橘品种以积累柠檬酸为主,极少数以积累苹果酸为主。本研究发现,黄果柑整个果实成熟期,柠檬酸含量占总酸含量的90%以上,说明黄果柑果实以积累柠檬酸为主。在柑橘花后6-12个月为成熟过程,常常伴随着明显的有机酸的降低[22-23]。本试验中,黄果柑果实有机酸含量在花后230或250 d开始逐渐降低,说明黄果柑果实有机酸的降解主要在果实成熟后期。成熟期黄果柑果实的总酸含量CK>红橘砧>枳壳砧>香橙砧,说明对黄果柑进行嫁接能够有效地降低果实有机酸含量,且香橙砧最好。
CS是一个变构酶,存在于果实细胞线粒体中,能催化草酰乙酸(OAA)与乙酰辅酶A(Ac-CoA)结合生成柠檬酸;PEPC能催化丙酮酸与CO2形成OAA,可供进一步合成柠檬酸;MDH能可逆催化苹果酸的分解与合成,对苹果酸的积累和柠檬酸的代谢具有重要作用。文涛等[24]研究发现CS活性与罗伯逊脐橙有机酸含量极显著正相关。本研究中,黄果柑果实CS活性在花后250 d之后,虽然有一定程度的降低,但是仍保持在一个较高的活性水平,而有机酸含量呈明显的下降趋势,可能是黄果柑有机酸含量的降低不取决于CS活性的降低,与CS活性关系不大,这与罗安才等[25]的试验结果一致。本试验中,不同砧木的PEPC活性随着黄果柑果实的成熟而不断降低,与龚荣高等[26]的研究一致,说明PEPC在黄果柑果实有机酸合成过程中起关键作用,其活性的降低能够抑制果实有机酸的合成。不同砧木间的CS、PEPC、MDH活性在各时期的差异性情况不同,说明其活性不是导致不同砧木间有机酸含量差异的主要因素。
顺乌头酸酶(ACO)是一个变构酶,催化柠檬酸向乌头酸的反应;异柠檬酸脱氢酶(IDH)与有机酸的合成、积累相关,果实中普遍存在NAD依赖型和NADP依赖型2种型式的IDH,线粒体中的NAD-IDH催化异柠檬酸的形成,细胞质中的NADP-IDH参与了柠檬酸的分解[27]。本试验中,黄果柑果实成熟后期,ACO活性缓慢下降,说明其活性与后期黄果柑果实有机酸快速下降关系不大,与Hirai等[18]在温州蜜柑上的研究结果一致。随着果实成熟,NADP-IDH的活性不断提高,与果实有机酸积累呈负相关,这与陈明[8]的结果一致。成熟后期,不同砧木间ACO和NADP-IDH活性差异显著,且表现为香橙砧>枳壳砧>红橘砧>CK,说明嫁接能够提高黄果柑果实ACO和NADP-IDH活性,从而促进有机酸的分解,ACO和NADP-IDH活性共同作用是导致砧木间有机酸含量出现差异的重要因素。
有机酸合成、代谢及分配受到许多因素控制,其中包括代谢酶相关基因的时空和组织特异性表达调控[28]。因此,明确不同砧木对有机酸代谢相关基因表达的影响在黄果柑砧木的选择中就显得十分必要。本研究发现,除花后270 d外,CS活性均表现为CK最高,说明嫁接能够抑制CS基因的表达,这可能是砧木影响黄果柑CS表达的细胞微环境的结果。除个别时期各个处理间PEPC和MDH相对表达量均无显著差异,说明砧木基本不影响黄果柑果实PEPC的基因表达,可能由黄果柑自身特性所决定,有待于进一步研究。枳壳砧果实的ACO表达峰值出现在花后230 d,CK和红橘砧的表达峰值出现在花后250 d,而香橙却在花后270 d,说明,在一定程度上,枳壳砧木提早了黄果柑有机酸的分解,香橙砧木延迟了有机酸的分解。除了花后210 d,砧木间NADP-IDH的表达量差异显著,且香橙砧>枳壳砧>红橘砧>CK,说明砧穗互作能够改变黄果柑果实NADP-IDH的表达。相关性分析结果表明:不同砧木的黄果柑NADP-IDH表达水平与总酸含量间呈显著性负相关,而CS、MDH、ACO与总酸含量均未表现出显著相关性。综上表明,NADP-IDH基因的表达可能是影响不同砧木黄果柑果实中有机酸含量的关键限制因子,这与王西成等[29]在葡萄上的研究结果相同。
[1]沈兆敏.中国柑桔区划与柑桔良种[M].北京:中国农业科技出版社,1988:192.
[2]汪志辉,刘世福,严巧巧,等.石棉县黄果柑生物学特性调查与差异株系比较[J].北方园艺,2011(14):20-24.
[3]Iglesias D J,Tadeo F R,Legaz F,et al.Invivosucrose stimulation of colour change in citrus fruit epicarps:Interactions between nutritional and hormonal signals[J].PhysiologiaPlantarum,2001,112(2):244-250.
[4]Chen Ming,Xie Xiulan,Lin Qiong,et al.Differential expression of organic acid Degradation-Related genes during fruit development of navel oranges (Citrussinensis) in two habitats[J].Plant Molecular Biology Reporter,2013,31(5):1131-1140.
[5]Zheng Q M,Tang Z,Xu Q,et al.Isolation,phylogenetic relationship and expression proling of sugar transporter genes in sweet orange(Citrussinensis)[J].Plant Cell Tiss Organ Cult,published online,2014,9,21.
[6]Terol J,Soler G,Talon M,et al.The aconitate hydratase family from Citrus[J].BMC Plant Biology,2010,10(1):58-59.
[7]关军锋.果实品质生理[M].北京:科学出版社,2008:15-16.
[8]陈明.椪柑和脐橙果实柠檬酸合成与降解相关基因表达及其调控研究[D].杭州:浙江大学,2013.
[9]郑永强,邓烈,何绍兰,等.几种砧木对哈姆林甜橙植株生长、产量及果实品质的影响[J].园艺学报,2010,37(4):532-538.
[10]淳长品.彭良志,雷霆,等.不同柑橘砧木对锦橙果实品质的影响[J].园艺学报,2010,37(6):991-996.
[11]刘翔宇,李娟,黄敏,等.柑橘砧木对砂糖橘果实糖积累的影响[J].中国农业科学,2015,48(11):2217-2228.
[12]杨义伶.柑橘砧木抗旱性评价及相关生理指标与基因表达差异分析[D].南昌:江西农业大学,2012.
[13]肖玉明.干旱胁迫对温州蜜柑果实品质及柠檬酸代谢相关基因表达的影响[D].长沙:湖南农业大学,2014.
[14]毛琼琼.南丰蜜橘果实糖酸代谢及其调控研究[D].南昌:江西农业大学,2013.
[15]Lin Qiong,Wang Chengyang,Dong Wencheng,et al.Transcriptome and metabolome analyses of sugar and organic acid metabolism in Ponkan (Citrusreticulata) fruit during fruit maturation[J].Gene,2015,554:64-74.
[16]Perotti V E,Del vecchio H A,Sansevich A,et al.Proteomic,metabalomic,and biochemical analysis of heat treated Valencia oranges during storage[J].Postharvest Biology and Technology,2011,62(2):97-114.
[17]郭燕.几个苹果品种果实糖酸积累及糖代谢相关酶活性变化研究[D].杨凌:西北农林科技大学,2012.
[18]Hirai M,Ueno I.Development of Critrus fruits:Fruits development and enzymatic changes in juice vesieletissue[J].Plant and Cell Physiology,1977,18:791-799.
[19]刘庆.暗柳甜橙红色突变体性状形成的分子机理研究[D].武汉:华中农业大学,2008.
[20]Zhang Bo,Shen Jiyuan,Wei Wenwen,et al.Expression of genes associated with aroma formation derived from the fatty acid pathway during peach fruit ripening[J].Journal of Agricultural and Food Chemistry,2010,58(10):6157-6165.
[21]Yamaki Y T.Organic acid in the jiuce ofcitrusfruit[J].Journal of the Japanese Society for Horticultural Sience,1989,58:587-594.
[22]Liu Yongzhong,Liu Qing,Xiong Jingjing,et al.Difference of a citrus late-ripening mutant(Citrussinensis)from its parental line in sugar and acid metabolism at the fruit ripening stage[J].Sci China SerC Life Sci,2007,50(4):511-517.
[23]Yu Keqin,Xu Qiang,Da Xinlei,et al.Transcriptome changes during fruit development and ripening of sweet orange (Citrussinensis)[J].BMC Genomics,2012,13:10.
[24]文涛,熊庆娥.柑桔果实糖、酸代谢研究概况[J].中国南方果树,2001,30(2):13-16.
[25]罗安才,杨晓红,邓英毅,等.柑橘果实发育过程中有机酸含量及相关代谢酶活性的变化[J].中国农业科学,2003,36(8):941-944.
[26]龚荣高,吕秀兰,张光伦,等.罗伯逊脐橙在不同生境下果实有机酸代谢相关酶的研究[J].果树学报,2006,23(6):805-808.
[27]蒲雪荔.石棉黄果柑增糖降酸效应研究[D].雅安:四川农业大学,2013.
[28]刘永忠,刘庆,熊晶晶,等.晚熟芽变脐橙在果实成熟阶段与原品种在糖酸代谢的差异[J].中国科学:C辑,2007,37(3):352-357.
[29]王西成,吴伟民,赵密珍,等.NAA对葡萄果实中糖酸含量及相关基因表达的影响[J].园艺学报,2015,34(3):425-434.
Effects of Different Rootstocks on Huangguogan Fruit Organic Acid Content,Acid Metabolism-related Enzyme Activity and Gene Expression
CAO Shuyan1,RONG Yi1,GU Xianjie1,LI Qingnan1,LIAO Ling1,YE Shuang1,QIU Xia1,WANG Zhihui1,2
(1.College of Horticulture,Sichuan Agricultural University,Chengdu611130,China;2.Institute of Pomology and Olericulture,Sichuan Agricultural University,Chengdu611130,China)
In order to explore the effects of different rootstocks on fruit organic acid,acid metabolism-related enzyme activities and the gene expression,to provide theoretical evidence for the selection of the rootstocks.In this experiment withPoncirustrifoliate(L.)Raf,CitrusreticulateBlanco,C.junos(sieb.) Tanaka for the rootstocks from Huangguogan seeding as the control for study.Showed that:Huangguogan fruit had been accumulated mainly citric acid,C.junos(sieb.) Tanaka could most effectively reduce the organic content of the fruit,mature period was 24.35% lower than in the the seedling.Grafting can be reduced to some extent the activity of CS,but has little effect on the activity of PEPC and MDH.250-330 days after flowering,the ACO and NADP-IDH activity exhibited a trend ofC.junos(sieb.) Tanaka>Poncirustrifoliate(L.)Raf>CitrusreticulateBlanco>CK.In addition to seeding,Huangguogan fruit between the level of expression ofNADP-IDHand total acid content was significantly negatively correlated,there was no significant correlation betweenCS,MDH,ACOand total acid content.TheC.junos(sieb.) Tanaka is ideal rootstock varieties of Huangguogan,Organic acid content of different rootstocks fruit is the result of combined action of the ACO and NADP-IDH activity,andNADP-IDHgene expression may be the mainly limiting factor of Huangguogan fruit organic acid with different rootstocks.
Huangguogan;Rootstock;Organic acid;Enzyme activity;Gene expression
2016-04-12
四川省科技支撑计划项目(2011NZ0034);四川省科技厅基金项目(10ZC1454)
曹淑燕(1991-),女,山东滨州人,在读硕士,主要从事果树栽培理论与技术研究。
汪志辉(1968-),男,四川眉山人,教授,博士,主要从事果树栽培理论与技术研究。
S666.9;Q78
A
1000-7091(2016)04-0080-08
10.7668/hbnxb.2016.04.014
