水分条件对红砂叶片碳同位素组成与光合特性和分枝生长的影响
2016-08-06胡小文
潘 佳,李 荣,胡小文
(兰州大学 草地农业科技学院草地农业生态系统国家重点实验室,兰州 730020)
水分条件对红砂叶片碳同位素组成与光合特性和分枝生长的影响
潘佳,李荣,胡小文*
(兰州大学 草地农业科技学院草地农业生态系统国家重点实验室,兰州 730020)
摘要:在盆栽条件下,研究了不同水分处理对红砂(Reaumuria soongorica)叶碳同位素组成、光合特性和分枝生长的影响,并进一步调查了自然条件下不同退化程度红砂草地的土壤含水量,分枝生长、叶碳同位素及其关系。结果表明:(1)在盆栽条件下,随土壤含水量的降低,红砂当年生分枝生物量、一级分枝长、二级分枝数及其叶片净光合速率、气孔导度和蒸腾速率均显著减小,而叶片碳同位素组成(δ13C)和水分利用效率则随土壤含水量降低而显著增加;且叶片δ13C与当年生分枝生物量、一级分枝长、二级分枝数、叶片净光合速率、气孔导度和蒸腾速率呈显著负相关关系。(2)在田间自然条件下,红砂叶片δ13C与立地30~60 cm及60~100 cm土层的土壤含水量、单位冠幅面积生物量、单位冠幅面积分枝数呈显著负相关关系。研究认为,在盆栽和田间条件下,红砂叶片δ13C是指示其生境水分状况的良好指标;红砂主要利用土壤的深层水分,其在土壤含水量相对较低的轻度退化区水分利用效率比土壤含水量相对较高的重度退化区更高。这一结论对于理解干旱生境中红砂的水分利用策略以及红砂草地的管理和恢复具有一定的指导意义。
关键词:红砂; 碳同位素组成;干旱胁迫;光合特性;水分利用效率
干旱胁迫是限制植物正常生长发育的主要因素,长期生长在干旱半干旱区的植物,通常对干旱生境具有一定的适应性,如一些旱生植物具有发达的根系以从土壤中获取充足的水分,而有些植物则可以通过生理调节来降低自身渗透势,进而促进水分吸收[1]。此外,在水分胁迫条件下,植物可通过降低气孔导度来控制水分的蒸腾速率,这一过程通常有利于植物保持较高的水分利用效率(water use efficiency, WUE),确保植物在干旱环境中正常生长[2]。
对植物个体而言,WUE是指在一段时间内干物质积累量和耗水量的比值;对植物叶片而言,是指在几秒和几分钟内净光合速率和蒸腾速率的比值[3],又称瞬时水分利用效率(instantaneous water use efficiency, WUEi)。由于WUE可客观反映植物对水分的利用状况及其对水分亏缺的耐受性,因而可作为特定环境选择合适物种的重要指标[4-5]。长期以来,围绕植物水分利用及其与环境、遗传的关系开展了大量的工作[4,6-7]。尤其是自碳同位素技术出现以来,δ13C的测算已经成为判断植物WUE的有效工具。δ13C部分取决于叶片胞间二氧化碳浓度与大气环境二氧化碳浓度的比值(Ci/Ca),它在一定程度上反映了CO2需求和供应的关系,而这种关系的变化主要由植物的光合能力以及气孔导度的改变而引起,这二者的改变间接决定了植物的WUE[5]。δ13C与WUE、植株生长以及水分胁迫生理反应之间的关系已有大量研究[3,6,8-9],但主要集中在农作物和乔木,而有关控制条件下荒漠灌木的研究则相对较少[10-11]。
红砂(Reaumuriasoongorica)属柽柳科,为超旱生多年生半灌木,荒漠草原优势种,主要分布在西伯利亚、中亚、非洲东部和中部以及中国等地区。在中国境内,从鄂尔多斯西部、阿拉善沙漠、河西走廊、北山、柴达木盆地到塔里木盆地和准噶尔盆地都有分布[12-13]。红砂具有很强的耐旱、耐盐性,对维持荒漠生态系统的稳定性和持续性具有重要作用。此外,红砂富含家畜和野生动物所需的养分,已成为当地的重要饲料作物,尤其是在干旱季节。由此可见,红砂具有较高的生态和社会价值[14]。但近些年来,由于气候干旱化以及人类活动,分布在中国西北荒漠草原的大多数红砂已严重退化或者呈现一定的退化趋势。因而,红砂草地的恢复重建和保育已成为当地农牧民以及相关政府部门亟待解决的一个重要生态议题。
植物种群对环境因子的反应主要表现在种群个体和个体构件上,如植株高度、分枝数目、枝条长度、冠幅面积等[15],在构件水平上研究种群和环境因子的关系,对揭示植物群落结构形成有重要意义[16]。前人研究[12-13,15]表明,红砂主要由含老叶的分枝组成,当年生分枝生长既反映了单株植物的生长条件又决定了下一季节植物的生长,因而红砂当年生分枝的生长是表征其生长状态的重要指标。一般认为,红砂立地条件的水分状况可能是决定其分枝生长的关键环境因子,也与其WUE密切相关,如当年降雨量增加1 mm,红砂叶片碳同位素组成降低0.01‰~0.015‰[11,17-18]。然而,在自然条件下测量碳同位素组成往往受到复杂环境条件的高度限制[11],如降雨量、坡度、草地退化状况等都可能影响到其微生境,进而使其生长与WUE呈现不同状态。基于此,本研究结合盆栽与田间试验,旨在确定:(1)干旱胁迫对红砂生长、光合作用和叶片碳同位素组成的影响;(2)叶片碳同位素组成与其光合作用、生长和土壤水分之间的关系;(3)自然条件下红砂的水分利用策略。这些信息将有助于认识和理解干旱环境中红砂的水分利用途径,进而应用于红砂草原的管理和恢复。
1材料和方法
1.1试验设计
1.1.1盆栽试验红砂种子于2002年11月采集于阿拉善荒漠草原(105°34′E,39°05′N,海拔1 360 m),干燥清选后于4 ℃条件下贮存待用。2003年6月,将红砂种子播种于育苗钵中,充分供水,室温条件(18~25 ℃)下培养。30 d后将大小一致的幼苗移栽到30 cm×26 cm(高度×直径)的花盆中并放置于兰州大学校园内(103°52′E,36°03′N,海拔1 517 m),每盆6株,共20盆。土壤基质采用砂土与粘土按照2∶1混合而成(砂土取自阿拉善荒漠,粘土取自兰州大学校园)。幼苗生长期间每2 d浇1次水,以确保供水充足,生长至次年5月5日进行干旱胁迫处理。干旱处理前,每花盆保留3株长势一致的植株。根据前期不同浇水管理下红砂的生长状况,试验设置4个水分处理梯度,分别为每隔4(W4)、8(W8)、12(W12)和16 d(W16)浇水1次,每次浇水量2.5 L(相当于47 mm降雨),试验期间各处理水分供应量分别为(包括降雨)720.2(W4)、444.2(W8)、303.2(W12)和256.2 mm(W16),每处理5个重复。其中,W4相当于充足供水条件,作为对照,而其它处理则相当于不同程度的水分胁迫。由于试验期间自然降雨量远远小于处理浇灌水量,其影响可以忽略不计,因而未采取避雨措施(图1)。
1.1.2田间试验田间试验样地位于阿拉善荒漠草原(105°34′E,39°05′N,海拔1 360 m)。该地区年平均气温8 ℃,年降雨量80~150 mm,主要降雨分布在夏季,年潜在蒸发量3 000~4 000 mm。该地区的优势种群是红砂和无芒隐子草(Cleistogenessongorica)。试验区域自1995年以来由于放牧,草地植被覆盖率和种类组成形成了以畜棚和饮水点距离为中心的辐射分布格局,总体上呈现离畜棚和饮水点距离越近,植被覆盖率越低,毒杂草比例越高的趋势。根据前人的研究,这种植被覆盖和组成上的差异,往往也体现在土壤含水量及其分布上[19]。基于此,本研究选取了离畜棚距离分别为30 m(重度退化)、300 m(中度退化)、800 m(轻度退化)处的9块25 m×20 m的围栏小区,每处理3块小区。

图1 2004年5月14日至7月9日兰州市降雨情况(日/月)Fig. 1 Rainfall in Lanzhou City during the experimental period of 14-May to 9-July 2004(day/month)
1.2测定指标及方法
1.2.1土壤含水量盆栽试验于2004年5月5日~7月11日期间,每10~20 d测定各处理土壤含水量1次。土壤含水量的测定采用自己设计的直径为1.5 cm的土钻自花盆中采取整个土层土壤后,80 ℃条件下烘干48 h至恒重,并计算土壤含水量。田间试验每年生长季时调查植被覆盖率和植被生产力,并用种子水分测定仪(503,CPN Corp. USA)监控0~100 cm土壤水分在生长季节的动态变化,每小区3个重复,每10 cm作为一个测量层。由于种子水分测定仪在测定浅层土壤含水量上的局限性,0~20 cm土层含水量采用土钻取土后,80 ℃条件下烘干48 h至恒重,并计算土壤含水量。
1.2.2枝条光合参数和叶面积在盆栽实验中于2004年6月14日上午9:00~11:00用便携式光合仪CI-301PS(CID,USA),测量红砂当年生二级分枝的净光合速率和蒸腾速率,每次选取植株上部南面朝向的当年生二级分枝进行测量,每株测定一个分枝,每分枝测定3次(即3次读数)。测量过的二级分枝用剪刀剪下后,采用叶面积仪(Model3100,LICOR U.S.A.)扫描测定其叶面积,进而计算光合速率。水分利用效率(WUE)以净光合速率与蒸腾速率的比值表示[20]。
1.2.3生长指标在盆栽试验中于2004年7月11日,剪下每株所有当年生分枝并按一级分枝和二级分枝分别计数(一级分枝为2003年从基茎上分出的枝条,二级分枝为一级分枝上再度分生出来的枝条);同时,每株分别随机选取10个一级分枝和二级分枝,用直尺测量其分枝长度;随后,将每株所有当年生分枝置于80 ℃条件下烘干48 h至恒重,测定生物量。在田间试验中,于2004年8月17日,在每个小区选取6株大小一致的植株,先测量冠幅和株高;后剪下所有当年生分枝并计数(所有单株均未见当年生二级分枝);然后,每株随机选取60个分枝来测量分枝长度并计算其平均值;最后,将所有分枝在80 ℃条件下烘干48 h至恒重,计算生物量。
1.2.4稳定碳同位素将盆栽和田间试验的干叶样本,分别用自来水冲洗干净,80 ℃烘干至恒重,研磨,过0.1 mm筛子。将处理好的样品送交兰州大学资源环境学院的稳定同位素实验室测定δ13C同位素值,测量精度0.05%。盆栽试验每个处理测5个重复,田间试验每个处理测6个重复。
1.3数据分析
数据多重比较分析采用Duncan法,数据显著性分析采用SPSS 19.0完成。不同的处理变量之间的关系用Pearson’s相关系数来计算确定。
2结果与分析
2.1不同干旱胁迫和退化程度下土壤含水量比较
盆栽试验不同处理间土壤含水量表现出一定差异,特别是在试验前期(5月20日至6月4日),所有处理间土壤含水量均表现出显著差异;而在试验后期,除W8与W12处理在土壤含水量上未表现出显著差异外,其他处理间土壤含水量均差异显著(图2)。整体而言,4种干旱胁迫处理之间形成了从湿润到干旱的水分梯度。
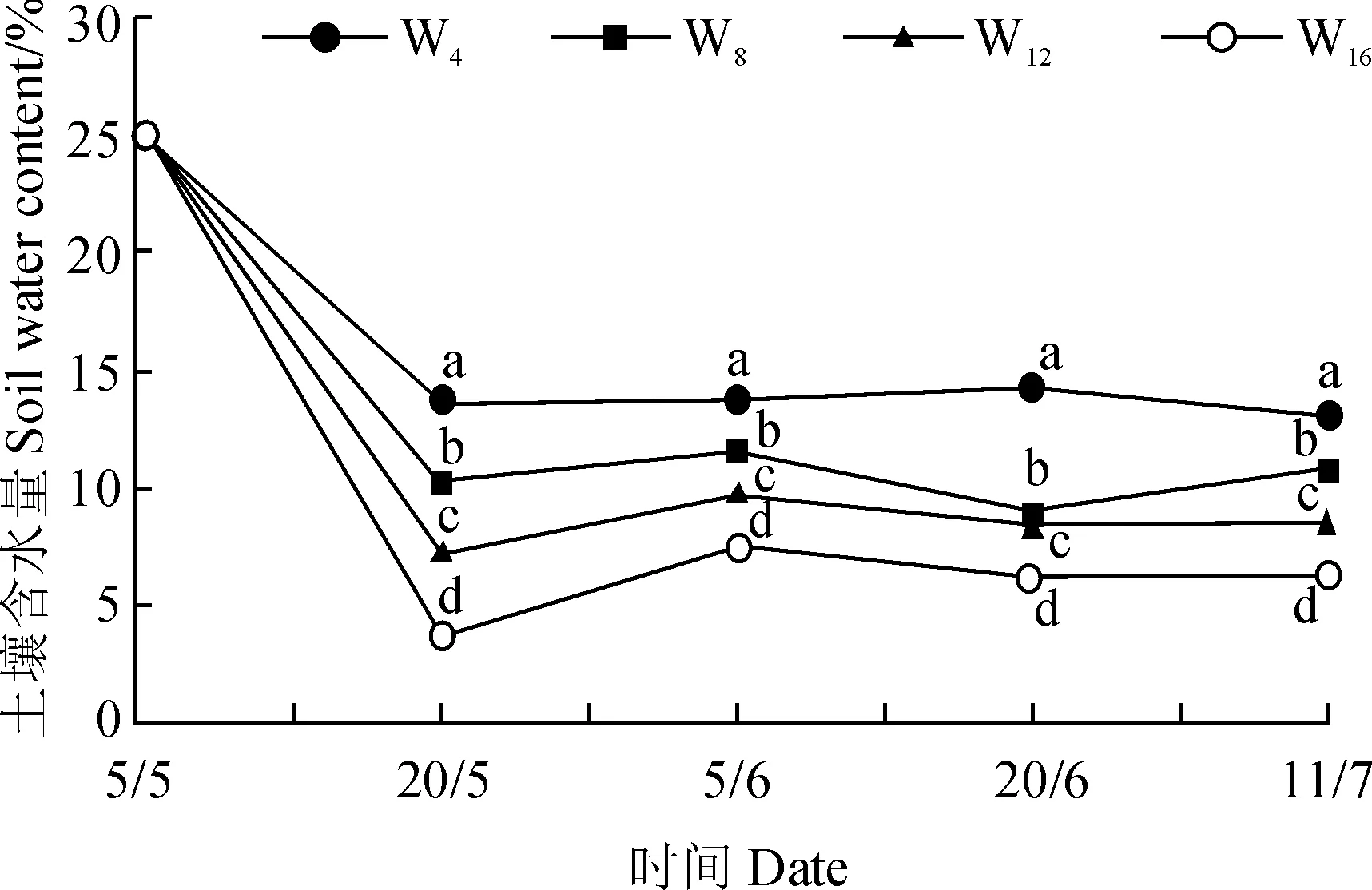
W4、W8、W12和W16分别表示每隔4、8、12、16 d浇1次水处理;同一时期不同字母表示处理间在0.05 水平存在显著性差异;下同图2 盆栽试验中不同供水条件下土壤含水量的动态变化W4,W8,W12 and W16 stand for the treatments irrigated every 4, 8, 12 or 16 days; The different letters at the same date indicate a significant difference among the treatments at 0.05 level; The same as belowFig. 2 The dynamics of soil water content of four potted treatments with different water supply
大田试验不同退化区间土壤含水量(SWC)随着土层深度的增加均表现出先增加后降低的趋势,各退化区土层土壤水分含量分布具有一定差异,尤其是轻度退化区(图3)。其中,0~30 cm土层土壤含水量(Swc30)在轻度退化区显著高于中度退化区和重度退化区(P<0.05);而40~60 cm土壤含水量(Swc60)在不同退化区之间差异不显著(P>0.05);70~100 cm土层土壤含水量(Swc100)却以轻度退化区最低,而以重度退化区最高,且轻度退化区与中度和高度退化区间差异显著(P<0.05),但中度和高度退化区间差异不显著(P>0.05)。
2.2不同干旱胁迫和退化程度对红砂分枝生长的影响
盆栽试验中,干旱胁迫显著影响了红砂植株高度、当年生分枝生物量、一级分枝长、二级分枝数和二级分枝长(P<0.05),但对一级分枝数的影响不显著(表1)。其中,在W16干旱胁迫处理下,红砂株高、当年生分枝生物量、一级分枝长和二级分枝数分别显著减少17.9%、59.2%、36.2%和79.7%,而二级分枝长则无显著变化。

图3 大田试验中不同程度退化区各土层土壤含水量Fig. 3 Soil water content (SWC) at each soil depth of different degraded sites
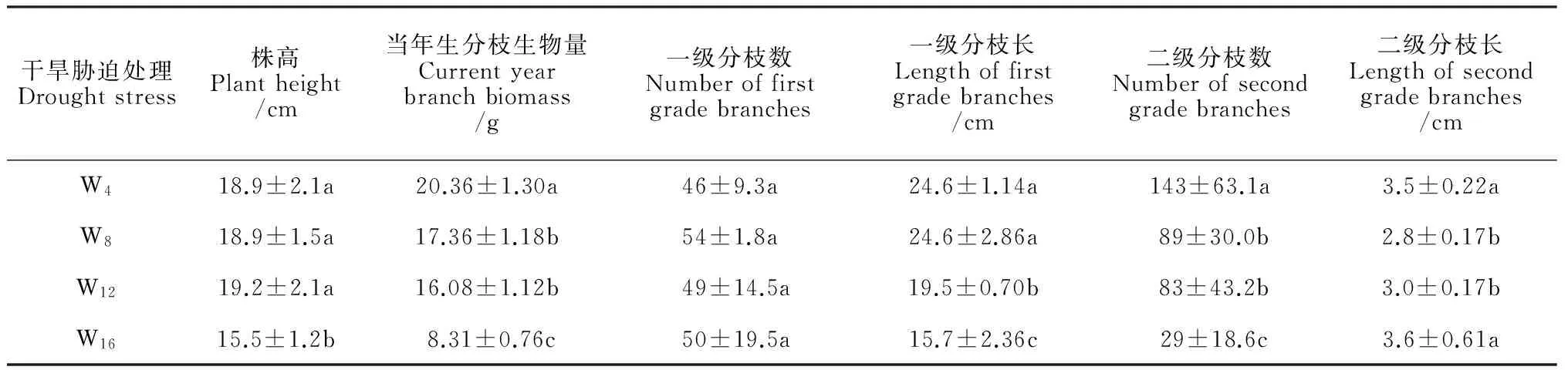
干旱胁迫处理Droughtstress株高Plantheight/cm当年生分枝生物量Currentyearbranchbiomass/g一级分枝数Numberoffirstgradebranches一级分枝长Lengthoffirstgradebranches/cm二级分枝数Numberofsecondgradebranches二级分枝长Lengthofsecondgradebranches/cmW418.9±2.1a20.36±1.30a46±9.3a24.6±1.14a143±63.1a3.5±0.22aW818.9±1.5a17.36±1.18b54±1.8a24.6±2.86a89±30.0b2.8±0.17bW1219.2±2.1a16.08±1.12b49±14.5a19.5±0.70b83±43.2b3.0±0.17bW1615.5±1.2b8.31±0.76c50±19.5a15.7±2.36c29±18.6c3.6±0.61a
注:同列不同字母表示处理间在0.05水平下存在显著性差异
Note: Numbers with different letters within the same column, are significant difference (P<0.05)
在大田试验(表2)中,从轻度退化区到重度退化区,红砂植株除一级分枝长、单位冠幅面积生物量呈增加趋势外,其他生长指标均呈不同程度的下降趋势,且大多达到显著水平(P<0.05)。其中,株高、一级分枝数和单位冠幅面积分枝数在不同退化区呈下降趋势且差异显著(P<0.05);红砂δ13C、单株的分枝生物量在不同退化区间呈下降趋势但无显著差异(P>0.05);与此相反,重度退化区红砂株高、单位面积冠幅生物量(BCA)明显高于轻度退化区和中度退化区(P<0.05)。
2.3干旱胁迫和退化程度对红砂光合特性和碳同位素比值的影响
在盆栽试验中(表3),随着干旱胁迫的加剧,红砂叶片净光合速率、蒸腾速率和气孔导度均呈逐渐显著下降的趋势,W16处理分别比W4处理显著降低75%、78%和83%(P<0.05);净光合速率在各干旱处理间均存在显著差异,而蒸腾速率、气孔导度在W12与W16处理之间没有显著差异。与上述三者变化不同,红砂叶片WUE随水分供应量的降低呈现先上升而后下降的趋势,并在W12处理下达到最高值,并显著高于其余处理。红砂叶片碳同位素(δ13C)组成值在盆栽试验中随干旱胁迫的加剧先增加后降低,并在W12和W16处理下显著高于W4与W8处理,而W12和W16处理间无显著差异(表3);在田间试验中,红砂叶片δ13C从轻度退化区到重度退化区基本呈下降趋势,但没有统计学差异(表2)。
2.4不同水分条件下红砂叶片δ13C与其生长、光合及土壤含水量间的关系
表4显示,盆栽试验的红砂叶片δ13C与其净光合速率、蒸腾速率、气孔导度、当年生分枝生物量、二级分枝数和二级分枝长之间呈极显著负相关(P<0.01),而与WUE呈显著正相关(P<0.05),与其余指标呈不显著负相关;同时,红砂叶片WUE与蒸腾速率和气孔导度呈显著负相关,而净光合速率与蒸腾速率、气孔导度、当年生分枝生物量、二级分枝数,蒸腾速率与气孔导度、当年生分枝生物量,以及气孔导度与当年生分枝生物量均呈显著正相关;另外,当年生分枝生物量与二级分枝数、一级分枝长呈极显著正相关,而一级分枝长和二级分枝数呈显著正相关,但二级分枝长和一级分枝数呈显著负相关。另外,从表5可以看出,田间试验的红砂叶片δ13C与单位冠幅面积分枝数、单位冠幅面积生物量、Swc60和Swc100呈显著负相关,但与冠幅面积呈显著正相关;此外,一级分枝数和株高与Swc30呈显著正相关,但与Swc60、Swc100呈显著负相关;而一级分枝长和单位面积冠幅生物量与Swc100呈显著正相关;一级分枝数和一级分枝长之间呈极显著负相关性。以上结果说明, 红砂植株的水分利用效率主要受到深层土壤水分的影响,分枝生长主要依赖土壤深层水分的供应。
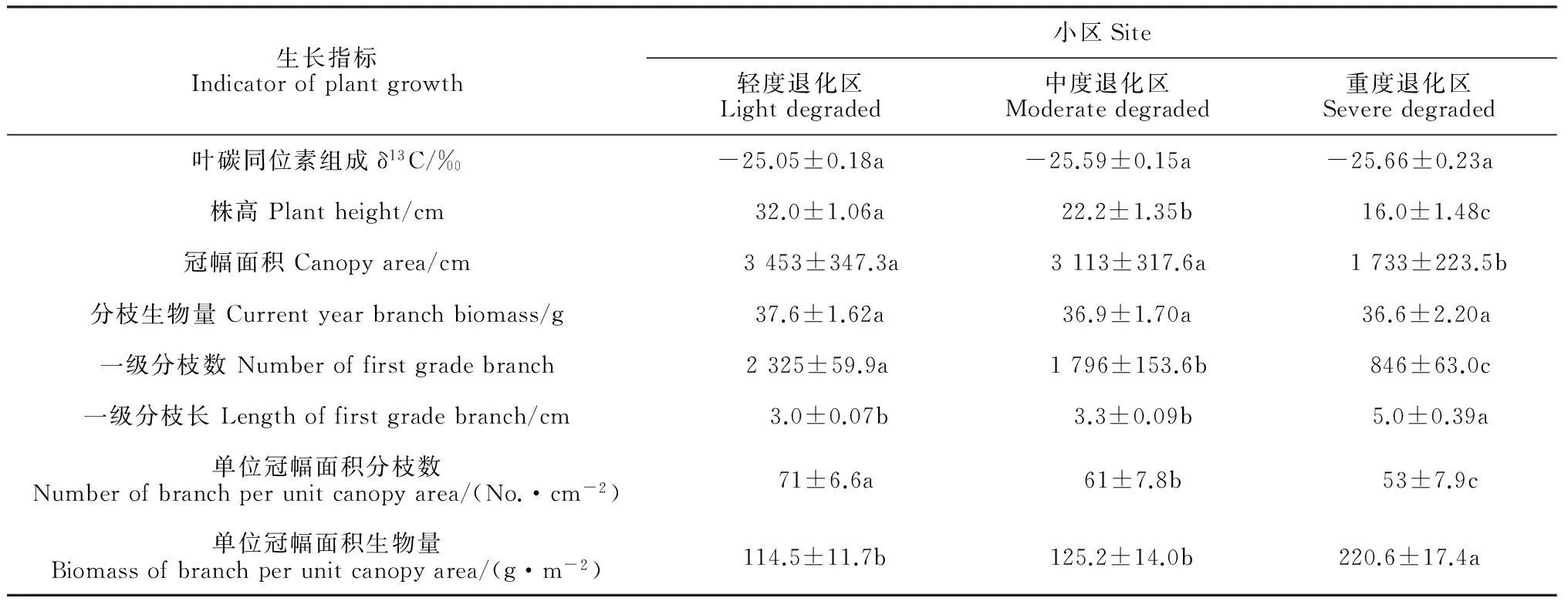
表2 不同退化程度草地红砂叶碳同位素组成及其生长特性
注: 同行不同字母表示处理间在0.05水平下存在显著性差异
Note: Different letters within a row indicate significant difference (P<0.05)

表3 干旱胁迫下红砂光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)、气孔导度(Gs)以及叶碳同位素(δ13C)组成
注: 同列不同字母表示处理间在0.05水平下存在显著性差异
Note: Numbers with different letters within the same column, are significantly different (P<0.05)

表4 干旱胁迫下红砂的形态学和生理学指标的相关性(盆栽试验)
注:δ13C 叶碳同位素组成;WUE.水分利用效率;Pn.净光合速率;Tr.蒸腾速率;Gs.气孔导度; DM.当年分枝生物量; NFG和LFG分别表示一级分枝数目和一级分枝长度; NSG和LSG分别表示二级分枝数目和二级分枝长度. *和**分别表示0.05和 0.01水平显著性相关关系;下同
Note:δ13C. Carbon isotope composition;WUE. Water use efficiency;Pn. Net photosynthesis;Tr. Transpiration rate;Gs. Stomatal conductance; DM. Current year branch biomass; NFG and LFG. Number and length of first grade branch, respectively; NSG and LSG. Number and length of second grade branch, respectively. * and ** indicate significant correlated at 0.05 and 0.01 level, respectively. The same as below

表5 植物生长特性、δ13C和土壤含水量的相关性(田间试验)
注: Swc30、Swc60、Swc100分别表示0~30、40~60、70~100 cm土层深度的土壤含水量; PH.株高;CA. 冠幅面积;NCA和BCA分别表示单位面积冠幅数和冠幅生物量
Note: Swc30, Swc60, Swc100 stand for soil water content at depth 0-30, 40-60, 70-100 cm, respectively; PH. Plant height; CA. Canopy area; NCA and BCA stand for number and biomass of branch per unit canopy area, respectively
3讨论
3.1盆栽条件下红砂光合特性及其分枝生长特征
在本研究中,从处理W4到W12红砂的净光合速率和蒸腾速率呈下降趋势,WUE呈上升趋势,这与前人在沙生灌木上的研究结论一致[21-24],即在干旱条件下最大净光合速率和蒸腾速率随水分降低而降低。干旱胁迫引起植物光合速率的降低可能由气孔或非气孔因素共同作用而致[25],如在本研究中,随干旱程度的加剧,气孔导度显著降低,气孔关闭或开度的降低在一定程度上会限制外源CO2的供应,从而引起光合速率的降低,这种情况下气孔限制往往是导致植物光合速率降低的主要原因。但同时我们也注意到,W12和W16处理在净光合速率上存在显著差异,但在气孔导度上二者却无明显差异,这表明在极度干旱胁迫条件下,除了气孔导度的变化外,其他机制如光合作用系统的同化能力、叶片的水分状况等非气孔因素也可能会影响到植物的光合速率[26-27]。如杨凤云[28]、冀宪领[29]等研究发现干旱胁迫使叶绿体的结构发生变化,植物膜受到损伤,膜脂过氧化加剧使产生超氧自由基,光合色素降解使光合电子传递系统遭到破坏,进而使合成酶活性下降、水解酶活性上升,从而导致光合速率下降。另外,红砂净光合速率与分枝生物量和二级分枝数呈显著正相关,可能意味着光合作用产生的同化物大部分被用于二级分枝的生长。虽然一级分枝长被认为是植物生长系统中最重要的部分[15],但本研究并没有发现净光合速率与一级分枝长及一级分枝数之间存在显著相关性。这可能是由于在测量光合之前,大多数一级分枝已经停止生长,与植物的分枝生长不同,因为光合作用参数在很大程度上只是植物瞬时生理状态的体现。
从盆栽和田间试验可以看出,从处理W4到处理W12,红砂叶片WUE和δ13C均呈增加趋势,且δ13C与WUE呈显著正相关,这一结果与邱权等[30]对青藏高原20种灌木幼苗WUE与叶片δ13C相关性的研究结果一致,因此用叶片δ13C表征植物WUE具有实际意义。这是因为,在干旱胁迫条件下,植物可通过降低气孔导度,减少蒸腾来提高其WUE[31]。但这一结论并不适用W16处理,因为与W12相比,W16处理的WUE和δ13C都降低了,这可能是由于严重干旱胁迫不仅影响气孔关闭,还对叶片光合系统造成损伤进而抑制生化反应(光合速率),从而在很大程度上降低了净光合速率和WUE[24-25]。此外还发现,红砂δ13C与干旱胁迫程度、净光合速率、当年生分枝生物量、二级分枝数、二级分枝长呈显著负相关,表明δ13C值在一定程度上可用于指示植物的生长性能及其所处水分条件[32]。
3.2不同退化区红砂的分枝生长和水分利用特征
在盆栽试验中,红砂叶片δ13C随干旱胁迫加剧而增加,且各处理间有显著差异,因此δ13C与干旱胁迫处理密切相关;而在田间试验中,红砂叶片δ13C与60和100 cm土层土壤含水量呈负相关,但与30 cm土层土壤含水量没有显著相关性,这间接地表明WUE主要受30 cm以下土壤含水量的影响,与先前的报道结论一致[33]。在其他灌木物种上也有一些类似的报道[34]。相反,Ma等[11]报道,红砂叶片的δ13C与30 cm深的土壤含水量显著相关,得出结论红砂主要利用的水分来自0~30 cm的土壤水。这在一定程度上可能是因为作者没有测量45 cm以下的土壤含水量,而可能得到一个不合适的结论。根据长期的调查研究,在阿拉善沙漠中30 cm以上的土壤含水量比较低,不能维持植物正常生长,但在60~100 cm深度土壤中有相对稳定的水位。另一个支持这个观点的结论是红砂发达的根系可以嵌入100 cm以下的土壤中[35]。但也有研究表明[36-37]在古尔班通古特沙漠南缘红砂为浅根植物,主要利用0~80 cm土壤水,而在黑河中游红砂可通过根系伸长生长利用185 cm以下土壤水分,因此在不同生境条件下红砂可调节根系形态特征来利用不同水源以适应其环境。Wang等[19]也发现,从轻度退化区到重度退化区红砂叶片WUE降低。本研究表明,从轻度退化区到重度退化区δ13C降低,但没有达到显著水平;与盆栽试验相比(叶碳同位素组成为-24.76 ‰~-27.37‰),田间试验的叶碳同位素组成(-25.05 ‰~-25.66‰)的变化要小得多。这可能是由于在盆栽试验中水分梯度差异更大,而在田间试验的3个小区中的水分条件差异相对较小;另一个原因可能是在有利的条件下成年期红砂比幼苗期红砂更容易保持水分平衡并有效地利用水分。显然,早围封的小区(如轻度退化区)更有利于植物高效利用水资源和提高植物产量。
3.3研究结果对红砂草地管理和恢复的启示
本研究表明,红砂株高、一级分枝数以及叶δ13C与100 cm深土层土壤含水量呈极显著负相关,表明红砂主要利用深层土壤水分;而从不同退化草地的水分分布状况来看,轻度退化区深层水分含量相对较低,因而相比重度退化区,其单位冠幅面积分枝数较少,生物量较低,这表明长期围封使轻度退化区红砂在土壤深层已形成了对水分的距离竞争,进而对其当年生生长造成了抑制作用,不利于植被的更新,可考虑适当加以利用来改善现状。而对于重度退化区,虽然其单株分枝生长较好,但其单株整体较为矮小,易被牲畜破坏,因而应继续围封来恢复其个体的健康生长。
参考文献:
[1]KOZLOWSKI T T, PALLARDY S G. Acclimation and adaptive responses of woody plants to environmental stresses[J].BotanicalReview, 2002, 68: 270-334.
[2]WHITEHEAD D, BEADLE C. Physiological regulation of productivity and water use in Eucalyptus: a review[J].forestEcologyandManagement, 2004, 193: 113-140.
[3]WRIGHT G C, HUBICK K T, FARQUHAR G D. Discrimination in carbon isotope of leaves correlated with water-use efficiency of field-grown eanut ultivars[J].AustralianJournalofPlantPhysiology,1988, 15, 815-825.
[4]吴英姿, 胡继超, 张雪松, 等. 环境因子对水稻叶片水分利用效率的影响[J]. 江苏农业科学, 2014, 42(5): 79-82.
WU Y Z, HU J CH, ZHANG X S,etal. Environmental factors influence water use efficiency on rice leaf[J].JiangsuAgriculturalSciences, 2014, 42(5): 79-82.
[5]FARQUHAR G D, O’LEARY M H, BERRY J A. On the relationship between carbon isotope discrimination and the intracellular carbon dioxide concentration in leaves[J].AustralianJournalofPlantPhysiology, 1982, 9: 121-137.
[6]LI C Y. Carbon isotope composition, water-use efficiency and biomass productivity ofEucalyptusmicrothecapopulations under different water supplies[J].PlantSoil, 1999, 214: 165-171.
[7]ZHANG X, WU N, LI C. Physiological and growth responses ofPopulusdavidianaecotypes to different soil water contents[J].JournalofAridEnvironments, 2005, 60: 567-579.
[8]陈平, 张劲松, 孟平, 等.稳定碳同位素测定水分利用效率----以决明子为例[J].生态学报, 2014, 34(19): 5 453-5 459.
CHEN P, ZHANG J S, MENG P,etal. Feasibility analysis on the determination of WUE by stable carbon isotope:CassiaobtusifoliaL. as an example[J].ActaEcologicaSinica, 2014, 34(19): 5 453-5 459.
[9]ZHENG S X, SHANGGUAN Z P. Spatial patterns of foliar stable carbon isotope compositions of C3plant species in the Loess Plateau of China[J].EcologicalResearch. 2007, 22: 342-353.
[10]LLORENS L, PENUELAS J, ESTIARTE M, Ecophysiological responses of two Mediterranean shrubs,EricamultifloraandGlobulariaalypum, to experimentally drier and warmer conditions[J].PhysiologiaPlantarum, 2003, 119: 231-243.
[11]Ma J Y, CHEN K., XIA D S. Variation in foliar stable carbon isotope among populations of a desert plant,Reaumuriasoongorica(Pall.) Maxim. in different environments[J].JournalofAridEnvironments, 2007, 69: 365-374.
[12]刘速, 刘晓云. 琵琶柴(Reaumuriasoongorica)地上植物量的估测模型[J].干旱区研究, 1996, 13: 36-41.
LIU S, LIU X Y, The estimating model of upper plant weight onReaumuriasoongoricasemi shrub[J].AridZoneResearch, 1996, 13: 36-41.
[13]MA M H, KONG L S. The bio-ecological characteristics ofReaumuriasoongoricaon the border of oasis at Hutubi, Xingjian[J].ActaPhytoecolgicaSinica, 1998, 22(3): 237-244.
[14]种培芳, 李毅, 苏世平, 等. 红砂3个地理种群的光合特性及其影响因素[J]. 生态学报, 2010, 30(4): 914-922.
ZHONG P F, LI Y, SU S P,etal. Photosynthetic characteristics and their effect factors ofReaumuriasoongoricathree geographical populations[J].ActaEcologicaSinica, 2010, 30(4): 914-922.
[15]徐莉, 王丽, 岳明, 等. 新疆阜康荒漠红砂种群构件结构与环境因子的灰色关联度分析[J]. 植物生态学报, 2003, 27(6):742-749.
XU L, WANG L, YUE M,etal. Analysis of grey relatedness between the modular structure ofReaumuriasoongoricapopulation in the desert offukang, xinjiang and the environmental factors[J].ActaPhytoecologicaSinica, 2003, 27(6): 742-748.
[16]黎云祥, 刘玉成, 钟章成. 植物种群生态学中的构件理论[M]. 生态学杂志, 1995, 14(6): 35-41.
LI Y X, LIU Y C, ZHONG Z C. Modular theory in plant population ecology[M].ChineseJournalofEcology, 1995, 14(6): 35-41.
[17]陈拓, 冯虎元, 徐世建, 等. 荒漠植物叶片碳同位素组成及其水分利用效率[J]. 中国沙漠, 2002, 22(3): 288-291.
CHEN T, FENG H Y, XU S J,etal. Stable carbon isotope composition of desert plant leaves and water use efficiency[J].JournalofDesertResearch, 2002, 22(3): 288-291.
[18]MA J Y, CHEN T, QIANG W Y,etal. Correlations between foliar stable carbon isotope composition and environmental factors in desert plantReaumuriasoongorica(Pall.) Maxim[J].JournalofIntegrativePlantBiology, 2005, 47(9), 1 065-1 073.
[19]王彦荣, 曾彦军, 张宝林, 等. 不同退化红砂荒漠草地的水分分配格局[J]. 应用生态学报, 2002, 13(8): 962-966.
WANG Y R, ZENG Y J, ZHANG B L,etal. Water distribution patterns in different degraded desert grassland ofReaumuriasoongorica[J].ChineseJournalofAppliedEcology, 2002, 13(8): 962-966.
[20]SINCLAIR T R, TANNER C B, BENNETT J M, Water-use efficiency in crop production[J].BiologyScienceAgronomySocietyofNewZealandSpecialPublication, 1984, 34: 36-40.
[21]FILELLA I, LIUSIA J, PIOL J,etal. Leaf gas exchange and the fluorescence ofPhillgrealatifolia,PistacialentiscusandQuercusilexsamplings in severe drought and high temperature conditions[J].EnvironmentalandExperimentalBotany, 1998, 39: 213-220.
[22]ZHENG W J, ZHENG X P, ZHANG C L. A survey of Photosynthetic carbon metabolism in 4 ecotypes ofPhragmitescommunisin northwest China: leaf anatomy, ultra-structure, and activities of ribulose 1, 5-bisphosphate carboxlase, phosphoenopyruvate carboxylase and glycollate oxidase[J].JournalofPlantPhysiology, 2000, 110: 201-208.
[23]韩刚, 赵忠. 不同土壤水分下4种沙生灌木的光合光响应特性[J]. 生态学报, 2010, 30(15): 4 019-4 026.
HAN G, ZHAO Z. Light response characteristics of photosynthesis of four xerophilous shrubs under different soil moistures[J].ActaEcologicaSinica, 2010, 30(15): 4 019-4 026.
[24]李文娆, 张岁岐, 丁圣彦, 等. 干旱胁迫下紫花苜蓿根系形态变化及与水分利用的关系[J]. 生态学报, 2010, 30(19): 5 140-5 150.
LI W R, ZHANG S Q, DING S Y,etal. Rootmorphological variation and water use in alfalfa under drought stress[J].ActaEcologicaSinica, 2010, 30(19): 5 140-5 150.
[25]范苏鲁, 苑兆和, 冯立娟, 等. 水分胁迫对大丽花光合作用、蒸腾和气孔导度的影响[J]. 中国农学通报, 2011, 27(8): 119-122.
FAN S L,WAN Z H,FENG L J,etal. Effects of water stress on photosynthesis, transpiration and stomatal conductance inDahlialeaves[J].ChineseAgriculturalScienceBulletin, 2011, 27(8): 119-122.
[26]EARL H J. Stomatal and non-stomatal restrictions to carbon assimilation in soybean (Glycinemax) lines differing in water use efficiency[J].EnvironmentalandExperimentalBotany, 2002, 48: 237-246.
[27]LIU M Z, JIANG G M, LI Y G,etal. Gas exchange, photochemical efficiency, and leaf water potential in threeSalixspecies[J].Photosynthetica, 2003, 41(3): 393-398.
[28]杨凤云. 土壤水分胁迫对梨树生理特性的影响[J]. 安徽农业, 2004, 6: 11-12.
YANG F Y. The influence of soil water stress on the pear tree physiological characteristics[J].AnhuiAgriculture, 2004, 6: 11-12.
[29]冀宪领, 盖英萍, 牟志美, 等. 干旱胁迫对桑树生理生化特性的影响[J]. 蚕业科学, 2004, 30(2): 117-122.
JI X L, GAI Y P, MOU Z M,etal. Effect of water stress on physiological and biochemical character of mulberry [J].ActaSericologicaSinica, 2004, 30(2): 117-122.
[30]邱权, 潘昕, 李吉跃. 青藏高原20种灌木幼苗生物量分配、水分利用效率及叶片δ13C比较[J]. 西北林学院学报, 2014, 29(4): 8-14.
QIU Q, PAN X, LI J Y. Comparison on biomass allocation and leaf water use efficiency and δ13C of 20 shurb seedlings in Tibetan platesu[J].JouralofNorthwestForestryUniversity, 2014, 29(4): 8-14.
[31]单立山, 李毅, 段雅楠, 等. 红砂幼苗根系形态特征和水分利用效率对土壤水分变化的响应[J].西北植物学报, 2014, 34(6): 1 198-1 205.
SHAN L S, LI Y, DUAN Y N,etal. Response of root morphology and water use efficiency ofReaumuriaSoongoricato soil water change[J].ActaBot.Boreal.-Occident.Sin, 2014, 34(6): 1 198-1 205.
[32]马剑英, 陈发虎, 夏敦胜, 等. 荒漠植物红砂稳定碳同位素组成的空间分布特征[J].第四纪研究, 2006, 26(6): 947-954.
MA J Y, CHEN F H, XIA D S,etal. Spatial distribution characteristics of stable carbon isotopecompo sitions in desert plantReaumuriasoongorica[J].QuaternarySciences, 2006, 26(6): 947-958.
[33]YODER C K, NOWAK R S. Soil moisture extraction by evergreen and drought-deciduous shrubs in the Mojave Desert during wet and dry years[J].JournalofAridEnvironments, 1999, 42: 81-96.
[34]DARROUZET-NARDI A, D’ANTONIO C M, DAWSON T E. Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow[J].PlantSoil, 2006, 285: 31-43.
[35]OLIVEIRA R S, BEZERRA L, DAVIDSON E A,etal. Deep root function in soil water dynamics in cerrado savannas of central Brazil[J].FunctionalEcology, 2005, 19: 574-581.
[36]XU H, LI Y. Water use strategy of three central Asian desert shrub and their response to rain pulse events[J].PlantandSoil, 2006, 285: 5-17.
[37]余绍文, 孙自永, 周爱国, 等. 用D、18O同位素确定黑河中游戈壁地区植物水分来源[J]. 中国沙漠, 2012, 32(3): 717-723.
YU S W, SUN Z Y, ZHOU A G,etal. Determination of water sources of Gobi plant by D and18O stable isotopes in middle reaches of the Heihe River[J].JournalofDesertResearch, 2012, 32(3): 717-723.
(编辑:裴阿卫)
文章编号:1000-4025(2016)06-1190-09
doi:10.7606/j.issn.1000-4025.2016.06.1190
收稿日期:2016-01-14;修改稿收到日期:2016-05-27
基金项目:农业部公益性行业科研专项(201403048-3);甘肃省重大科技攻关项目(2013 GS 05907)
作者简介:潘佳(1991-),女,在读硕士研究生,主要从事作物栽培与耕作研究。E-mail:panj14@lzu.edu.cn *通信作者:胡小文,教授,硕士生导师,主要从事草业科学、作物栽培与育种研究。E-mail:huxw@lzu.edu.cn
中图分类号:Q945.79
文献标志码:A
Effect of Water Conditions on Carbon Isotope Composition, Photosynthesis and Branch Growth ofReaumuriasoongorica
PAN Jia, LI Rong, HU Xiaowen*
(College of Pastoral Agriculture Science and Technology, Lanzhou university, State Key Laboratory of Grassland Agro-ecosystems, Lanzhou 730020,China)
Abstract:Present study determined the effect of soil moisture condition on leaf carbon isotope composition (δ13C), photosynthesis and current year branch growth of Reaumuria soongorica in the pot experiment. The branch growth and δ13C of R. soongorica growing in different degraded area were investigated, and their relationship with soil moisture content that plant stand were analyzed. The results showed that:(1) The current year branch biomass (DM), length of first grade branches (LFG), number of second grade branches (NSG), net photosynthesis rate (Pn), stomatal conductance (Gs) and transpiration rate (Tr) of R. soongorica were largely reduced as water supplies decreased in the pot experiments. δ13C and water use efficiency(WUE) increased as water supplies reduced, and a significant negative correlation was observed between δ13C and DM,LFG,NSG,Pn,Gs and Tr. (2) In the field, δ13C decreased from light degraded sites to severe degraded one and significantly negatively correlated with soil water content at a depth of 30-60 cm and 60-100 cm. A negative relationship was also observed between δ13C and biomass per unit canopy area (BCA) and number of first grade branch per unit canopy area (NCA). These results suggested that δ13C is a good indicator of R. soongorica response to water stress both under potted and field conditions and R. soongorica mainly utilize water in deeper soil and utilize water more efficiently in light degraded area than that in severe degraded area. This conclusion may improve understanding water use strategies of R. soongorica in the arid environments, and have practical use in grassland management and restoration.
Key words:Reaumuria soongorica; carbon isotope composition; drought stress; photosynthesis; water use efficiency
