外源亚精胺对鸭梨自花授粉花粉管伸长及花柱蛋白表达的影响
2016-03-18李中勇尹宝颖徐继忠
岳 雷,李中勇,尹宝颖,张 媛,徐继忠
(1.河北农业大学 园艺学院,河北 保定 071001;2.沧州市农林科学院,河北 沧州 061001)
外源亚精胺对鸭梨自花授粉花粉管伸长及花柱蛋白表达的影响
岳雷1,2,李中勇1,尹宝颖1,张媛1,徐继忠1
(1.河北农业大学 园艺学院,河北 保定071001;2.沧州市农林科学院,河北 沧州061001)
摘要:为探讨外源多胺调控鸭梨自交不亲和的生理生化机制,分别以自花授粉和外源亚精胺处理后1~3 d的花柱为试材,测定了不同处理不同时期花粉管生长发育动态,利用双向电泳技术和质谱分析技术分析了外源亚精胺处理前后不同时期鸭梨花柱蛋白的差异表达。结果显示:与对照相比,不同浓度的Spd处理均能极显著促进花粉管生长,0.25 mmol/L Spd处理72 h后可使大量花粉管到达花柱基部;在自交花柱电泳图谱中发现了3种特异蛋白质,A蛋白质MrA=28.0 kDa,pI=5.7;B蛋白质MrB=28.5 kDa,pI=5.7;C蛋白质MrC=22.0 kDa,pI=5.3,这3种特异蛋白质可能与自交不亲和性相关;质谱分析获得了2个较好的蛋白点,分别为推定和假定蛋白。
关键词:亚精胺;鸭梨;花粉管;花柱蛋白
梨属植物(PyrusL.)为配子体型自交不亲和性果树,绝大多数品种自花授粉不结实。自花授粉后,花粉管在沿花柱向子房生长途中受到抑制而停止生长是不能完成授粉受精的主要原因[1]。徐义流等[2]发现,丰水梨授粉后36 h,自花授粉的花粉管仅伸长到花柱全长的44.3%位置,并停止生长,而异花授粉的花粉管已达到花柱的基部。在配子体自交不亲和研究中需要解决的关键问题是花柱如何识别自己和异己的花粉管,解决此问题的着眼点集中在分离S位点的基因及其产物上。自证实配子体自交不亲和性花柱抑制花粉管生长的物质主要是S-RNase后,许多果树的S基因所对应的蛋白质被分离和鉴定[3-5]。
多胺是一类小分子生物活性物质,在植物细胞生长和分裂中起重要作用,参与植物花芽分化、花和果实的发育及各种生理胁迫反应。已有研究表明,多胺与果树花粉的萌发和花粉管的伸长密切相关[6-10],适宜浓度外源多胺可促进梨[6]、苹果[7]、猕猴桃[8]自花授粉后花粉管的伸长。但目前关于多胺与自交不亲和关系的研究多集中于花粉管生长发育表型[6-8]及内源激素含量测定[9,11-12]等生理指标上,而对授粉受精过程中外源多胺对花柱内相关蛋白的表达缺乏深入研究。本研究以自交不亲和品种鸭梨为试材,探讨外源亚精胺处理对鸭梨自交授粉花柱内相关蛋白表达的影响,并进行差异点质谱分析,以期为揭示外源多胺调控鸭梨自交结实的生理生化机制提供理论依据。
1材料和方法
1.1试验材料
于花前采取鸭梨树冠外围带饱满花芽的健壮枝条,带回实验室置于光照培养箱内培养,开花后分别进行人工自花和喷施亚精胺后自花授粉处理,以不同处理1~3 d的花柱为试验材料。
1.2试验方法
1.2.1花粉液体培养花柱提取液参考齐国辉[13]方法略加修改。取各处理花柱0.1 g,加1 mL预冷的基本培养液(10%蔗糖,0.01%硼酸)冰浴研磨成匀浆,以12 000 r/min离心20 min,上清液再用基本培养液稀释1倍即为花柱提取液。在洗净的双凹玻片上滴上花柱提取液,将花粉均匀点播在上面,放入(25±1)℃的培养箱保湿避光培养12 h,每个处理观察3个视野,重复4次。拍照并用Spot图像分析软件统计不少于100粒花粉粒和20个花粉管,数据经处理后用Duncan′s新复极差法进行多重比较。
1.2.2花粉管生长的动态观察在授粉后24,48,72 h采取物候期一致的第1,2朵边花,每次5朵,去掉花瓣和萼片后放入装有FAA固定液(70%酒精∶甲醛∶冰醋酸=90∶5∶5)的小瓶中,抽气固定后备用。固定的花柱用清水冲洗后,用8 mol/L的氢氧化钠软化10 h,清水冲洗后放入事先配置好的过夜的脱色苯胺蓝溶液(0.1%苯胺蓝+0.1 mol/L磷酸钾)中染色1 h(阴暗处),然后将花柱和子房分开,花柱直接在载玻片上用盖玻片压片,用ZEISS Axioskop 40型荧光显微镜观察花粉管生长并拍照。
1.2.3可溶性蛋白的提取参照张金锐等[14]方法略加修改。称取30 mg样品,加入0.6 mL蛋白提取缓冲液(62.5 mmol/L Tris-HCl pH值6.8,20 g/L SDS,15 g/L DTT),4 ℃下研磨并完全移至1.5 mL离心管中,在4 ℃下提取1 h后,于15 000 r/min离心20 min,取上清液置入干净离心管,按4∶1(V/V)加入变性样品缓冲液(60 mmol/L Tris-HCl pH值6.8,25%甘油,2%SDS,5%β-巯基乙醇,0.1%溴酚蓝),沸水浴8 min后备用。
SDS不连续电泳过程参照汪家政等[15]的方法(垂直板不连续电泳),分离胶浓度为12%,浓缩胶浓度为5%,电极缓冲液为Tris-甘氨酸(pH值8.3),含0.1%SDS。点样量15 μL,浓缩胶稳压80 V,分离胶稳压160 V,至溴酚蓝指示剂距胶底1 cm时结束电泳。
考染显色的凝胶用BIO-RAD凝胶成像仪拍照,用Quantity One分析软件对图像进行分析。
1.2.4蛋白质双向电泳样品制备参照何瑞锋等[16]的方法并加以改进。取一定量样品放入液氮中研成粉末,随后加入-20 ℃预冷的10%三氯乙酸(丙酮配制,内含有0.07%β-巯基乙醇)-20 ℃沉淀2 h。于4 ℃,13 000 r/min条件下离心20 min。弃上清,加入冷丙酮(其中含0.07%β-巯基乙醇)中,-20 ℃放置1 h(期间每隔一定时间振荡一次使色素充分脱去),同上离心。重复2~3次。将沉淀悬浮于80%冷丙酮中-20 ℃放置1 h,离心去丙酮后将沉淀于-20 ℃放置,待丙酮挥发干净即可备用。使用前按每mg干粉加20 μL样品溶解液(9.5 mol/L尿素,4% NP-40,2.5%两性电解质载体(其中0.5% pH值3.0~9.5,1.0% pH值4.0~6.0,1.0% pH值6.0~9.0),5%β-巯基乙醇),37 ℃保温30 min以上,上样前于15 000 r/min离心20 min,上清液上样或-70 ℃保存。
采用IEF/SDS-PAGE双向电泳,参照O′Farrell[17]的方法,略有改动。第一向电泳采用微型等电聚焦柱状电泳,凝胶长9 cm,直径2 mm。凝胶组成为:3.78%丙烯酰胺,0.22%甲叉双丙烯酰胺,9.0 mol/L尿素,2% NP-40,3% Ampholine pH值 3.5~10.0。负极电极液为20 mmol/L NaOH,正极电极液为10 mmol/L H3PO4,加样(每管15 μL),进行电泳50 V×1 h,100 V×30 min,250 V×30 min,500 V×6~7 h。聚焦电泳结束后,剥下胶柱放入平衡液(60 mmol/L Tris-HCl pH值6.8,3%SDS,5%β-巯基乙醇,10%甘油,微量1%溴酚蓝)中,30 ℃平衡25~30 min。第二向为SDS-PAGE微型平板电泳,分离胶浓度为12%,浓缩胶浓度为5%。将固定后的第一向胶柱横放在第二向凝胶板的上端,并在近胶柱碱性端一侧加入标准蛋白质标记液10 μL,再用融化的1%琼脂糖将胶柱封固,然后进行第二向电泳,稳流20 mA/板,大约2 h 30 min结束。用银染法染色,拍照记录。
1.2.5差异点质谱分析根据总蛋白2-DE图谱上显示的差异蛋白点,将蛋白点A~C从凝胶上切取后送北京华大蛋白质研发中心有限公司,进行肽质指纹图谱测定,查询与结果相匹配的蛋白质,并将氨基酸序列在NCBI上搜索,根据同源的序列,推测其功能。
2结果与分析
2.1外源亚精胺对鸭梨花粉管伸长的影响
试验结果表明(表1),不同浓度的Spd处理后花柱提取液都能促进花粉管生长。授粉后1 d,0.50 mmol/L Spd处理的花柱提取液花粉管长度为261.8 μm,极显著高于对照和其他3种浓度Spd处理水平。授粉后3 d,花粉管长度以0.25 mmol/L Spd处理的效果最好,为对照长度的123.35%,差异达极显著水平。

表1 Spd处理花柱提取液对花粉管生长的影响
注:同一列中不同小写字母表示差异显著(P<0.05);不同大写字母表示差异极显著(P<0.01)。
Note:Different lowercase in the same column represent significant difference(P<0.05);Different capital letters represent extremely significant difference(P<0.01)。
对鸭梨花柱进行人工自花授粉,通过荧光观察(图1-3),授粉24 h后,花粉刚在柱头萌发,48 h后可看到花粉管伸进到花柱1/4处,72 h后花粉管在花柱中部停止伸长,花粉管先端膨大呈球状。0.25 mmol/L Spd处理鸭梨花柱后自花授粉,进行荧光观察(图4-6)可以看出,授粉24 h后花粉在柱头大量萌发且花粉管有一定伸长,48 h后花粉管伸长并穿过花柱中部,72 h后大量花粉管到达花柱基部。
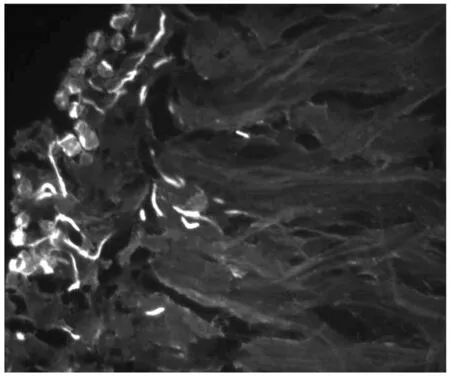
图1 自花授粉24 h(柱头)
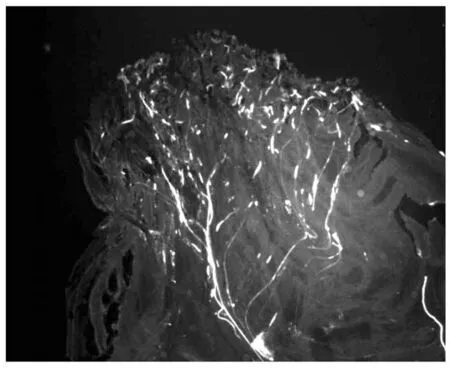
图2 自花授粉48 h(上部)

图3 自花授粉72 h(花粉管末端)

图4 Spd处理授粉后24 h(上部)
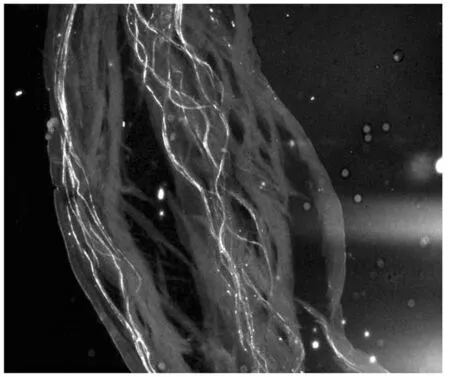
图5 Spd处理授粉后48 h(中部)
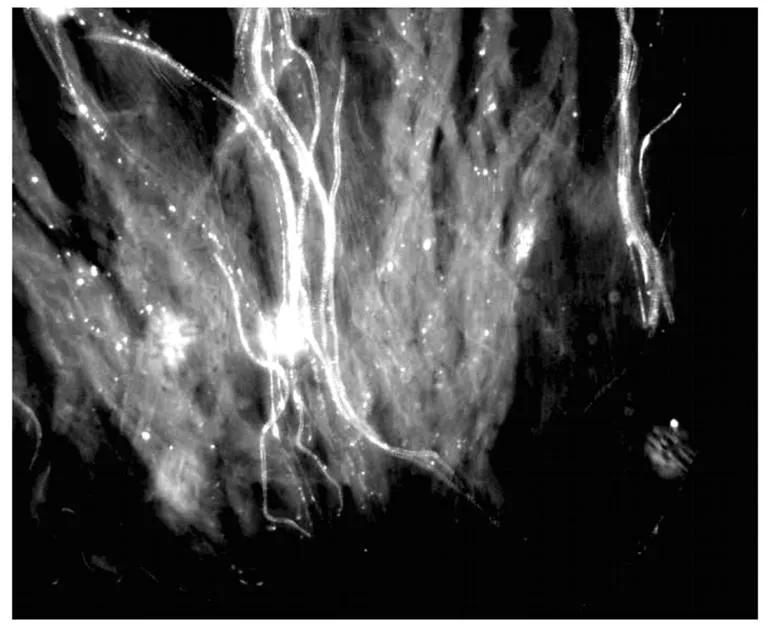
图6 Spd处理授粉后72 h(基部)
2.2自花授粉和亚精胺处理后鸭梨花柱可溶性蛋白表达的比较
分别取鸭梨自花授粉和0.25 mmol/L Spd处理后的鸭梨花柱,提取可溶性蛋白进行SDS-PAGE分析,由图7可以看出,在自花授粉1,2 d的花柱中,存在一条分子量为25.5 kDa的蛋白带(6泳道右侧箭头所示),这条带在自花授粉后3 d的花柱中消失。
对0.25 mmol/L Spd处理后的鸭梨花柱提取可溶性蛋白进行SDS-PAGE分析,由图7可以看出,分子量为24.5,32.5 kDa的蛋白带只存在于Spd处理后1 d的花柱中,而在Spd处理后2,3 d的花柱中没有表达(2泳道右侧箭头所示)。分子量为30.0 kDa的蛋白带存在于Spd处理后1,2 d的花柱中,而在3 d的花柱中没有表达(3泳道左侧箭头所示)。
对比自花授粉和0.25 mmol/L Spd处理后的鸭梨花柱单向电泳蛋白条带,发现分子量为18.0 kDa的蛋白带在自花授粉花柱中均有表达(7泳道右侧箭头所示),而其只在Spd处理后3 d的花柱中微弱表达。分子量分别为22.5,30.0 kDa的2条蛋白带在自花授粉3 d花柱中表达(7泳道左侧箭头所示),而在Spd处理后3 d花柱中没有表达。

图7 自花授粉和Spd处理后鸭梨花柱蛋白比较
2.3自花授粉和亚精胺处理后鸭梨花柱蛋白的双向电泳分析
对比自花授粉的鸭梨花柱和0.25 mmol/L Spd处理后鸭梨花柱的双向电泳图谱可以看出(图8-13),蛋白点主要分布在等电点(pI)4.0~8.0,分子量(Mw)20.0~100.0 kDa。应用ImageMaster 2D Platinum软件对2块胶进行分析,将凝胶去除纹理和消除背景,进行自动点检测,手工消除因杂质或明显的瑕疵形成的“假点”,添加或切割因距离过近被识别成同一蛋白的蛋白点,每块胶图都能得到约200个蛋白点。然后利用软件进行差异点分析,结合肉眼观察,在矩形区域(分子量:21.0~35.0 kDa、等点电:pI 4.5~6.5)差异比较明显。

图8 鸭梨自花授粉1 d花柱蛋白双向电泳图谱

图9 Spd处理后1 d鸭梨花柱蛋白双向电泳图谱
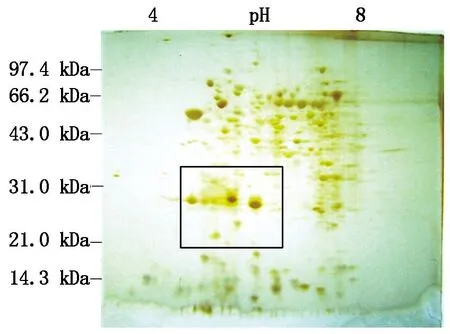
图10 鸭梨自花授粉2 d花柱蛋白双向电泳图谱
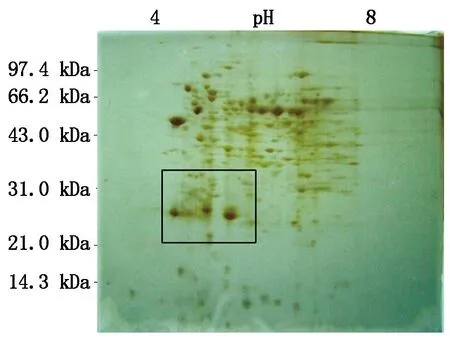
图11 Spd处理后2 d鸭梨花柱蛋白双向电泳图谱
对比0.25 mmol/L Spd处理后鸭梨花柱与自花授粉的鸭梨花柱双向电泳图谱(图14)可以看出,蛋白点A(28.0 kDa,pI=5.7)存在于自花授粉1~3 d的花柱中,而在Spd处理的花柱中,其表达量随授粉时间延长而减弱。蛋白点B(28.5 kDa,pI=5.7)存在于自花授粉1~3 d的花柱中,而该点在Spd处理花柱中,随授粉时间的延长,其表达量逐渐减弱并消失。蛋白点C(22.0 kDa,pI=5.3)存在于自花授粉1~3 d和Spd处理1 d的花柱中,而在Spd处理后2,3 d的花柱中没有表达。

图13 Spd处理后3 d鸭梨花柱蛋白双向电泳图谱

1-3.鸭梨自花授粉1~3 d的花柱;4-6.Spd处理后1~3 d的鸭梨花柱。
综上所述,本试验可以确定这个区域(分子量约为21.0~35.0 kDa、等点电pI 4.5~6.5)的蛋白与鸭梨自交不亲和性相关,同时也能说明在鸭梨花期喷施外源多胺,能够影响授粉受精过程中花柱内蛋白质表达的差异。
2.4差异蛋白点的初级质谱结果分析
根据上述确定的差异蛋白,挖取了部分分辨率良好的差异蛋白质点,在北京华大蛋白质研发中心进行了肽指纹图谱分析。对蛋白点A~C进行了MALDI-TOF-MS分析,将所得的结果在http://www.matrixscience.com/数据库中进行查询,得到2个较好的蛋白匹配结果(表2)。
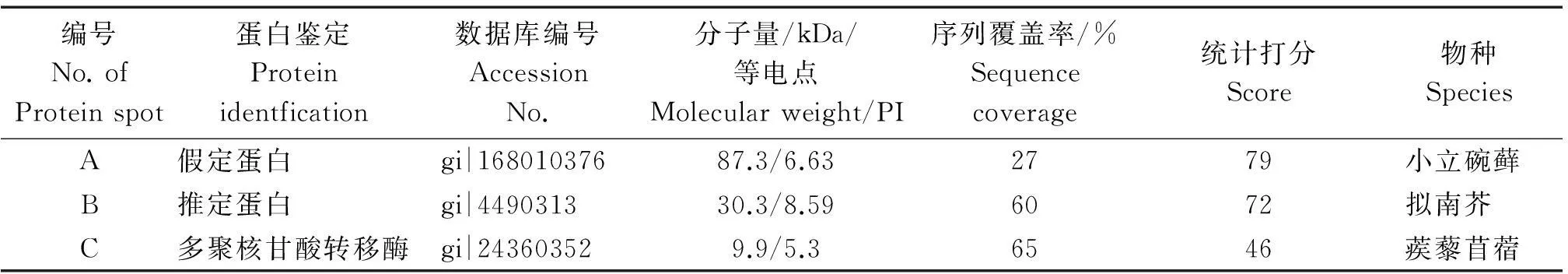
表2 通过MALDI-TOF质谱分析和PMF检索确定的蛋白点
3讨论与结论
陈迪新等[18]研究认为,适宜浓度的多胺可以促进丰水梨花粉萌发和花粉管生长。徐继忠等[19]研究表明,喷施3种多胺均能够加速苹果花粉管的伸长,从而缩短授粉受精时间,提高授粉受精能力。本试验研究结果表明,亚精胺处理后花柱提取液对鸭梨花粉管生长的影响与浓度和处理天数有关,通过荧光观察,0.25 mmol/L亚精胺处理能够促进活体鸭梨花粉管生长,3 d后使其穿过花柱中部到达花柱基部。
配子体型自交不亲和果树之所以表现为自交不亲和,可能是由于花粉管在花柱生长过程中,花柱内S复等位基因产物——具RNase活性的S糖蛋白在花柱内分解同源花粉管的RNA,而抑制花粉管生长所造成的[20-21]。陈学好等[22]对黄瓜子房施用外源Spd处理,在花后1~3 d的子房中出现了4种特异的蛋白质,从而影响单性结实和子房(幼果)发育。肖华山等[23]对荔枝花芽分化过程中多胺、RNA/DNA以及蛋白质/RNA比值动态的进行研究,揭示了多胺在DNA复制、转录和翻译3个水平上都能影响生物大分子的合成,参与了蛋白质合成的调节。本试验通过SDS-PAGE分析发现,与自花授粉花柱相比,0.25 mmol/L Spd处理后3 d的鸭梨花柱有2条蛋白带(22.5,30.0 kDa)没有表达。Spd处理后不同时期的花柱蛋白表达也存在明显差异,在处理后1 d的花柱中有2条特异蛋白带(24.5,32.5 kDa)表达,分子量为30.0 kDa的蛋白带在处理后3 d的花柱中消失。这可能与外施多胺通过影响核酸的构象及各种酶的活性而影响蛋白质的合成或降解有关[24]。细胞蛋白质的表达和功能行为在部分程度上能通过翻译后的降解来互补调控,有目的地降解细胞内蛋白质是许多细胞程序调控的重要模式[3]。本研究通过双向电泳技术对上述处理进一步蛋白分离,发现在Spd处理的花柱中,蛋白点A随着授粉天数的延长,其表达量减弱。蛋白点B随着授粉天数的延长,其表达量减弱直至消失。蛋白点C存在于自花授粉1~3 d和Spd处理后1 d的花柱中,而在Spd处理后2,3 d的花柱中消失。这些变化可能是由于多胺能刺激蛋白质合成的各个中间步骤,如激活一系列氨酰_tRNA合成酶与核糖体结合,起装备核糖体和稳定亚基的作用,并参与蛋白质合成的起始、延长、终止各个过程[25],从而降低花柱内具RNase活性的S糖蛋白浓度,打破鸭梨自交不亲和性。
本试验选取了3个蛋白点进行肽质指纹分析,通过数据库检索,得到2个有意义的结果。蛋白点A~B的结果匹配第一的结果分别为假定蛋白和推定蛋白,蛋白功能尚未知晓;蛋白点C的匹配结果分数较低,但其功能与影响核糖核酸酶有关。
参考文献:
[1]Zhang S L,Hiratsuka S.Analysis varietal differences in self-and cross-incompatibility reactions of Japanese pears using stylar culture technique[J].Journal of the Japanese Society for Horticultural Science,1999,68:373-383
[2]徐义流,张绍铃.以~(32)P示踪观察梨花粉管在自花与异花花柱中生长的方法[J].植物生理学通讯,2003,39(3):238-239.
[3]李天忠,张志宏.现代果树生物学[M].北京:科学出版社,2008.
[4]GU Qing-qing,Zhang Q L,HU Hong-ju.Identification of self-Incompatibility genotypes in some sand pears (PyruspyrifoliaNakai) by PCR-RFLP analysis[J].Agricultural Sciences in China,2009,8(2):154-160.
[5]姜新,曹晓艳,王大江,等.南疆杏品种自交不亲和S-RNase基因型的鉴定[J].果树学报,2012,29(4):569-576.
[6]齐国辉,徐继忠,邵建柱,等.多胺、MGBG、水杨酸对鸭梨和雪花梨花粉萌发及花粉管生长的影响[J].河北农业大学学报,2008,31(3):21-27.
[7]张雪梅,李保国,齐国辉,等.外源多胺对不同苹果品种自花授粉花粉管生长的影响[J].南方农业学报,2012,43(9):1344-1348.
[8]齐秀娟,张绍铃,方金豹.植物生长调节剂对猕猴桃花粉萌发的影响[J].经济林研究,2010,28(3):45-50.
[9]吕文君,郭素娟,邹峰,等.板栗授粉前后子房中内源多胺含量的变化[J].西北林学院学报,2013,28(5):109-114.
[10]高兴颖,崔世茂,付崇毅,等.外源亚精胺对日光温室砂糖橘生殖生长的影响[J].沈阳农业大学学报,2013,44(5):704-707.
[11]徐继忠,陈海江,马宝焜,等.外源多胺对富士苹果花和幼果内源多胺与激素的影响[J].园艺学报,2001,28(3):206-210.
[12]李六林,张绍铃,郭艳玲.新高梨花粉败育与内源多胺含量变化的关系[J].园艺学报,2007,34(2):301-304.
[13]齐国辉.鸭梨自交不亲和与亲和变异的生理生化特性及分子机理研究[D].保定:河北农业大学,2005.
[14]张金锐,刘勇,林刚,等.SDS-PAGE分析转基因小麦与主栽小麦杂交后代的高分子量麦谷蛋白亚基[J].生物技术通讯,2006,17(3):341-344.
[15]汪家政,范明.蛋白质技术手册[M].北京:科学出版社,2000.
[16]何瑞锋,丁毅,张剑锋,等.植物叶片蛋白质双向电泳技术的改进与优化[J].遗传,2000,22(5):319-321.
[17]O′Farrell P H.High resolution two dimentional electrophoresis of proteins[J].J Biol Chem,1975,250:4007-4021.
[18]陈迪新,张绍铃.多胺及其合成抑制剂对梨花粉萌发及花粉管生长的影响[J].果树学报,2002,19(6):377-380.
[19]徐继忠,史常青,章文才.外源多胺对苹果花粉管伸长及胚珠寿命的影响[J].园艺学报,1998,25(1):92-93.
[20]张绍铃,房经贵,杨记磙.果树自交不亲和性的遗传与生理机制及其研究[J].果树学报,2001,18(1):49-52.
[21]Hiratsuka S,Kitoh Y,Matsushima J.Induction of deformed pollen tube tips and their morphological characterisitics in self-incompatible Japanese pear[J].Japan Soc Hort Sci,1991,60:257-265.
[22]陈学好,于杰,徐强,等.Spd和MGBG对黄瓜子房内源多胺和蛋白质组成的影响及与单性结实的关系[J].园艺学报,2005,32(4):632-637.
[23]肖华山,吕柳新,陈志彤.荔枝花芽分化过程中多胺、核酸和蛋白质的动态[J].应用与环境生物学报,2006,12(5):640-642.
[24]Rimpy K,Chowhan,Laishram R.Polyamines in modulating protein aggregation[J].Journal of Protein and Proteomics,2012,3(2):141-150.
[25]Tassoni A,Buuren M V,Fraceschetti M,et al.Polyamine content and metabolism inArabidopsisthalianaand effect of spermidine on plant development[J].Plant Physiology Biochemistry,2000,38(5):383-393.
《华北农学报》征订启事
《华北农学报》1986年创刊,由河北、北京、天津、河南、山西、内蒙古六省市区农科院、农学会联合主办,为全国首家跨省、市、区多单位联办的农业学术刊物。本刊立足华北,面向全国和全世界。主要刊载农业基础学科学术论文、研究报告及科研简报,报道农业学术动态。主要服务于农业高等院校师生和农业科研机构的研究人员。
《华北农学报》为中国科学引文数据库核心期刊(CSCD核心库)、中文核心期刊、中国科技核心期刊、RCCSE中国权威学术期刊(A+)和中国农业核心期刊。在2014年版《中文核心期刊要目总览》综合性农业科学类核心期刊中排名第2位,为我国有影响力的农业学术刊物。《华北农学报》多次荣获国家级及省级奖励:全国优秀科技期刊评比三等奖、全国优秀农业期刊学术类一等奖、首届华北优秀期刊、首届北方十佳期刊、中国北方优秀期刊、河北省优秀期刊、河北省十佳期刊及河北省荣誉期刊等奖项;2011年被评选为“中国精品科技期刊”。
《华北农学报》国内外公开发行, 国内统一刊号:CN13-1101/S,国际刊号ISSN 1000-7091。双月刊,双月28日出版,国际标准大16开本,240页,每期定价12元,全年72.00元。邮发代号: 18-10,国外发行代号:5918。全国各地邮局均可订阅。可随时汇款到编辑部订阅,请注明刊名、份数、姓名、地址、邮编及电话。
欢迎订阅、欢迎投稿。
地址:石家庄市和平西路598号《华北农学报》编辑部
邮编:050051
电话:0311-87652166
E-mail:hbnxb@163.com
网址:http://www.hbnxb.net/(投稿请登录学报网址注册投稿)
Effects of Exogenous Spd on the Pollen Tube Growth and Stylar Protein Expression During Pollination and Fertilization of Yali Pear
YUE Lei1,2,LI Zhongyong1,YIN Baoying1,ZHANG Yuan1,XU Jizhong1
(1.College of Horticulture,Agricultural University of Hebei,Baoding071001,China;2.Cangzhou Academy of Agriculture and Forestry Sciences,Cangzhou061001,China)
Abstract:Yali pear belongs to the kind of gametophytic self-incompatibility,the percentage of self-pollinated fruit set is extremely low.Our previous research showed that sprayed polyamine in a concentration range at florescence could break self-incompatibility of Yali pear.In order to explore the physiological and biochemical mechanism,the effects of exogenous Spd on the pollen tube growth and stylar protein expression of the Yali pear were studied by two-dimensional gel electrophoresis and mass spectrum analysis technology.The main results were as follows:The pollen tube length were extremely significantly increased after suitable concentration of Spd treatment,the best effects of exogenous Spd were 0.25 mmol/L Spd treatment,the number of pollen tube reached to the bottom of style were the most after 72 hours of treatment.Three specific proteins were found in the electrophoretogram.Two protein spots were obtained by the mass spectrometry results,which were predicted protein and putative protein respectively.
Key words:Spermidine;Yali pear;Pollen tube;Stylar protein
doi:10.7668/hbnxb.2016.01.004
中图分类号:Q78;S661.03
文献标识码:A
文章编号:1000-7091(2016)01-0022-07
作者简介:岳雷(1982-),男,河北沧州人,助理研究员,硕士,主要从事果树结实生理与分子生物学研究。通讯作者:徐继忠(1964-),男,河北唐山人,教授,博士,主要从事果树结实生理与分子生物学研究。
基金项目:河北省自然科学基金资助项目(C200600429)
收稿日期:2015-12-12
