益生菌与动物肠道自由基的关系
2014-12-21王进波初佳丽齐莉莉杨在宾
王进波 初佳丽, 齐莉莉 杨在宾
(1.浙江大学宁波理工学院,宁波 315100;2.山东农业大学,泰安 271018)
肠道是物质与能量代谢十分旺盛的器官,氧化还原反应剧烈,细胞代谢产生大量自由基。一定水平的自由基对肠道免疫功能具有积极作用,但过量自由基会引起肠道损伤、脂质过氧化、加速细胞衰老等一系列危害。肠道益生菌对动物营养物质的消化吸收、肠道组织发育、免疫调节及抑制致病菌的侵染等有着重要生理作用。益生菌调节动物肠道生理功能的机制十分复杂,最近的研究发现,益生菌可通过调控肠道自由基水平发挥重要生理作用。本文对益生菌在通过调控肠道自由基水平,既可发挥免疫调节作用,又防止对肠道氧化损伤的功能及其相关机制进行了综述。
1 细菌诱导肠道组织产生自由基的机制
肠黏膜上皮组织及细胞受到某些细菌刺激后,会产生活性氧自由基(ROS),这些 ROS参与肠道生理功能的调控。细菌诱导的肠道细胞ROS主要是由还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶家族(NADPH oxidase,NOX)催化合成的,NOX家族中NOX1和双氧化酶(dual oxidase,DUOX)在肠道组织中表达丰度很高[1]。病原菌能够刺激机体肠道组织产生并释放大量ROS,这些ROS可以协助机体对抗病原菌侵染,同时,也会引起严重的炎症反应,造成肠黏膜损伤[2-3]。病原菌释放游离的尿嘧啶碱基作为信号分子,通过G蛋白偶联受体(G protein-coupled receptors,GPCRs),激活肠黏膜上皮细胞中的DUOX,诱导合成大量 ROS(图 1)[3-5]。与病原菌不同,部分益生菌也能刺激肠道产生少量的ROS,这些ROS不仅不会损伤肠道,还对肠道维持正常的生理功能有益。Alam等[6]研究发现,益生菌能够通过刺激肠道吞噬细胞膜上的甲酰肽受体(formyl peptide receptors,FPRs),激活 NOX2,诱导机体肠道吞噬细胞产生 ROS。Leoni等[7]的研究指出,乳酸菌诱导的细胞ROS产生及细胞增殖依赖于肠上皮细胞的NOX1。Jones等[8]利用从果蝇肠道中分离的乳酸杆菌进行的研究表明,该菌能够通过激活NOX1诱导肠道组织产生释放ROS,这些ROS可以刺激小鼠、果蝇等模型动物肠道干细胞的分化,促进肠组织更新。Yan等[9]的研究则发现,益生菌能够通过产生可溶性蛋白成分,诱导宿主肠道合成一定量的ROS,有助于肠道细胞的存活及其生长增殖。益生菌的诱导肠道细胞产生ROS的机制与致病菌完全不同,其通过细胞壁上肽聚糖分子中的N-甲酰肽作用于肠黏膜细胞膜上的FPRs,激活细胞中的 NOX1,合成一定量的ROS(主要是超氧阴离子),这些ROS可以激活相关信号通路,刺激肠黏膜组织的更新(图1)[3]。

图1 细菌诱导肠黏膜上皮细胞产生ROS的机制Fig.1 The mechanism of ROS released from intestinal epithelial cells induced by the bacteria[3]
2 益生菌对肠道自由基的清除作用
过量的自由基会引起肠道组织受损,肠黏膜通透性增加,进而诱发肠黏膜溃疡和炎症反应;对肠道尚未发育完全的动物而言,自由基造成的肠黏膜损伤会导致其死亡[10-12]。肠道益生菌参与清除机体肠道代谢产生的自由基,从而保护肠道组织免于自由基的氧化损伤。益生菌产生的过氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)、含锰假性过氧化氢酶以及硫氧还蛋白还原酶、烟酰胺腺嘌呤二核苷酸氧化酶等还原酶具有清除ROS的作用[13-16]。肠道中的ROS主要包括超氧阴离子、过氧化氢(H2O2)和羟基自由基等,其中羟自由基的活性最强,对细胞核酸、蛋白质和脂类等均有很强的氧化性,其能够显著诱导益生菌抗氧化相关酶的表达,激发益生菌的抗氧化能力[17]。
Kullisaar等[17]从1名健康儿童的肠道菌群中分离得到2株抗氧化菌株,这2株菌对过氧化氢、超氧阴离子、羟自由基等活性氧具有明显的抗性。韩伟等[18]从腌制蔬菜中分离的5株菌均表现较强的自由基清除能力,这些菌株对1,1-二苯基-2-三硝基苯肼(DPPH)、羟自由基等的清除率和脂质过氧化的抑制率相对较高。白明等[19]研究了乳杆菌、双歧杆菌、嗜热链球菌和乳酸乳球菌等40株益生菌的抗氧化能力,结果证明,多数益生菌具有清除自由基的能力,但种属和菌株间差别较大,其中乳酸乳球菌清除自由基能力最强,嗜热链球菌次之,乳杆菌、双歧杆菌则比较弱。这些体外试验结果表明,益生菌能够清除自由基,其主要机制可能是自由基刺激了菌体相关还原酶系的活性,从而激发了益生菌的还原能力,进而清除自由基。
体内试验也表明,益生菌可提高动物机体的抗氧化能力,清除体内代谢产生的自由基,发挥抗衰老作用。不同研究者以小鼠为模型,研究凝结芽孢杆菌、双歧杆菌、德氏乳杆菌、富硒沼泽红假单胞菌等益生菌的抗氧化作用,结果表明,其血清、肝脏、脑等组织中SOD、GSH-Px等相关还原酶的活性提高,丙二醛(MDA)含量下降,抗衰老相关基因的表达水平上调[20-26]。闻平等[27]的研究发现,明串珠菌可提高家兔血清GSH-Px、SOD等还原酶的活性,降低血清MDA含量,并维持较长时间,这提示明串珠菌及其发酵产物有提高家兔抗氧化能力的作用。这些体内研究结果提示,益生菌能够刺激机体还原酶的表达或提高其活性,从而协助机体清除代谢产生的ROS,从而发挥抗氧化作用。
3 自由基在益生菌发挥生理功能过程中的作用
ROS的产生是吞噬细胞对细菌(包括致病菌和共生菌)免疫应答的重要特征,这提示ROS在益生菌与宿主互作过程中发挥重要作用。肠道益生菌能够诱导果蝇肠道组织氧化酶系统的表达,刺激肠道细胞产生过氧化氢,进而抑制病原菌在肠道中的增殖及其侵染作用,从而维持肠上皮的完整性[28-29];植物体受到细菌刺激时也会产生ROS,从而抑制病原菌对自身的侵染,保护自身组织、细胞免受病原破坏[2]。Kumar等[1]的研究发现,鼠李糖乳酸杆菌(Lactobacillus rhamnosus GG,LGG)能够显著刺激Caco-2细胞释放ROS,这些ROS能够诱导细胞泛素-蛋白酶体信号途径中泛素连接酶 12(ubiquitin conjugating enzyme 12,Ubc12)半胱氨酸残基的氧化失活,进而抑制核转录因子-κB(NF-κB)炎症信号通路的活性,从而发挥其免疫调节作用。Lin等[30]利用小鼠进行的体内试验也得出相似的结论,LGG诱导的ROS能够导致细胞内Ubc12失活,抑制泛素-蛋白酶体信号途径活性,进而阻止NF-κB炎症信号通路激活,阻止细胞炎症反应,并保护幼龄动物肠道黏膜组织的完整性,防止腹泻等疾病的发生。
此外,ROS还作为炎症因子及生长因子刺激的细胞信号转导过程的第二信使。在这一过程中,益生菌诱导产生的ROS会导致细胞信号通路相关蛋白的巯基迅速氧化失活,从而发挥第二信使功能。细胞内的酪氨酸磷酸酶就是这样一类受ROS调节的巯基蛋白酶,而酪氨酸磷酸酶是丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、黏附斑激酶(focal adhesion kinase,FAK)和NF-κB信号通路的重要调节因子,这些信号通路在机体肠道免疫调节、细胞增殖和运动等过程中发挥着重要作用(图2)[31-32]。因此,肠道益生菌可通过刺激机体产生ROS,调节上述细胞信号通路,从而发挥相关生理调节作用。
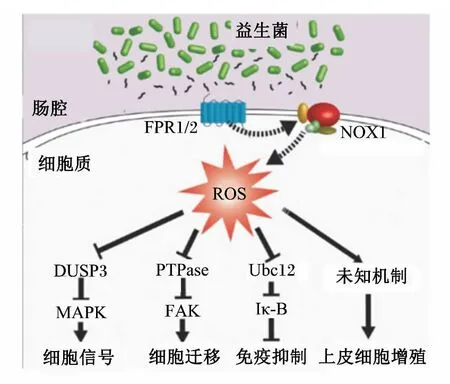
图2 益生菌诱导的ROS在细胞信号转导中的作用Fig.2 The roles of ROS induced by probiotics in cell signaling transduction [33]
4 小结
受到细菌等刺激后,产生释放自由基是一种十分保守的免疫调节机制,从植物到高等动物均存在类似的机制。动物肠道中生存着大量微生物,这些微生物通过调节肠道细胞自由基的水平,影响相关细胞信号通路的活性,从而调节机体肠道生理功能,这是肠道益生菌发挥生理作用的重要机制。解析益生菌诱导及清除肠道自由基的机制,揭示自由基作为第二信使在益生菌刺激的细胞信号转导过程中的作用,对全面了解肠道益生菌的免疫调节作用机理、探究肠道菌群与宿主健康的关系以及新型益生菌制剂的开发具有重要意义。但益生菌通过何种菌体成分、以何种互作方式激活肠道细胞信号通路诱导ROS的产生?益生菌参与调节维持肠道ROS的稳态的机制如何?这一系列问题仍需要进一步研究。
[1] KUMAR A,WU H,COLLIER-HYAMS L S,et al.Commensal bacteria modulate cullin-dependent signaling via generation of reactive oxygen species[J].The EMBO Journal,2007,26(21):4457-4466.
[2] KIM S H,LEE W J.Role of DUOX in gut inflammation:lessons from Drosophila model of gut-microbiota interactions[J].Frontiers in Cellular and Infection Microbiology,2013,3:116.
[3] PATEL P H,MALDERA J A,EDGAR B A.Stimulating cROSstalk between commensal bacteria and intestinal stem cells[J].The EMBO Journal,2013,32(23):3009-3010.
[4] LEE K A,KIM S H,KIM E K,et al.Bacterial-derived uracil as a modulator of mucosal immunity and gut-microbe homeostasis in Drosophila [J].Cell,2013,153(4):797-811.
[5] LEE K A,LEE W J.Drosophila as a model for intestinal dysbiosis and chronic inflammatory diseases[J].Developmental& Comparative Immunology,2014,42(1):102-110.
[6] ALAM A,LEONI G,WENTWORTH C C,et al.Redox signaling regulates commensal-mediated mucosal homeostasis and restitution and requires formyl peptide receptor 1[J].Mucosal Immunology,2014,7(3):645-655.
[7] LEONI G,ALAM A,NEUMANN P A,et al.Annexin A1,formyl peptide receptor,and NOX1 orchestrate epithelial repair[J].The Journal of Clinical Investigation,2013,123(1):443-454.
[8] JONES R M,LUO L,ARDITA C S,et al.Symbiotic lactobacilli stimulate gut epithelial proliferation via Nox-mediated generation of reactive oxygen species[J].The EMBO Journal,2013,32(23):3017-3028.
[9] YAN F,CAO H,COVER T L,et al.Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth[J].Gastroenterology,2007,132(2):562-575.
[10] TOMITA T,SADAKATA H,TAMURA M,et al.Indomethacin-induced generation of reactive oxygen species leads to epithelial cell injury before the formation of intestinal lesions in mice[J].Journal of Physiology and Pharmacology,2014,65(3):435-440.
[11] PERRONE S,TATARANNO M L,SANTACROCE A,et al.The role of oxidative stress on necrotizing enterocolitis in very low birth weight infants[J].Current Pediatric Reviews,2014,10(3):202-207.
[12] PERRONE S,TATARANNO M L,STAZZONI G,et al.Oxidative injury in neonatal erythrocytes[J].The Journal of Maternal-Fetal& Neonatal Medicine,2012,25(Suppl.5):104-108.
[13] BRUNO-BÁRCENA J M,ANDRUS J M,LIBBY S L,et al.Expression of a heterologous manganese superoxide dismutase gene in intestinal lactobacilli provides protection against hydrogen peroxide toxicity[J].Applied and Environmental Microbiology,2004,70(8):4702-4710.
[14] ROCHAT T,GRATADOUX JJ,GRUSSA,et al.Production of a heterologous nonheme catalase by Lactobacillus casei:an efficient tool for removal of H2O2and protection of Lactobacillus bulgaricus from oxidative stress in milk[J].Applied and Environmental Microbiology,2006,72(8):5143-5149.
[15] SERRANO L M,MOLENAAR D,WELS M,et al.Thioredoxin reductase is a key factor in the oxidative stress response of Lactobacillus plantarum WCFS1[J].Microbial Cell Factories,2007,6(1):29.
[16] TALWALKAR A,KAILASAPATHY K,HOURIGAN J,et al.An improved method for the determination of NADH oxidase in the presence of NADH peroxidase in lactic acid bacteria[J].Journal of Microbiological Methods,2003,52(3):333-339.
[17] KULLISAAR T,ZILMER M,MIKELSAAR M,et al.Two antioxidative lactobacilli strains as promising probiotics[J].International Journal of Food Microbiology,2002,72(3):215-224.
[18] 韩伟,刘文群,黄丽婵,等.5株微生物抗氧化作用的初步研究[J].食品与机械,2008,24(5):45-47,51.
[19] 白明,孟祥晨.益生菌抗氧化活性及菌体抗氧化相关成分的分析[J].食品与发酵工业,2009,35(5):6-11.
[20] 刘锋,张娟娟,张海,等.凝结芽孢杆菌TBC169株对亚急性衰老模型小鼠的抗衰老作用[J].中国老年学杂志,2009,29(4):433-436.
[21] 王跃,张宏娟,刘明方,等.双歧杆菌脂磷壁酸抗氧化作用的实验研究[J].中国老年学杂志,2007,27(13):1254-1257.
[22] 陈施羽.小白鼠肠道菌群中乳酸菌的筛选及其抗衰老作用初探[D].硕士学位论文.成都:四川师范大学,2007:1-66.
[23] 王挺,李弋.双歧杆菌对D-半乳糖致衰老小鼠的抗衰老作用[J].中国老年学杂志,2007,27(1):45-48.
[24] AKYOL S,MAS M R,COMERT B,et al.The effect of antibiotic and probiotic combination therapy on secondary pancreatic infections and oxidative stress pa-rameters in experimental acute necrotizing pancreatitis[J].Pancreas,2003,26(4):363-367.
[25] 曩潇潇,何瑶,王晓炜,等.富硒沼泽红假单胞菌对D-半乳糖致衰老小鼠抗氧化系统的影响[J].安徽农业科学,2007,35(16):4850-4852.
[26] 龚灵芝,陈小连,徐建雄.微生物源性抗氧化剂对高不饱和脂肪酸饲料致大鼠自由基损伤模型的影响[J].饲料工业,2008,29(20):32-34.
[27] 闻平,黄锡全,汪毅,等.L.SP.HXQ001 菌对家免抗氧化能力的影响[J].中国微生态学杂志,2002,14(2):76.
[28] HA E M,OH C T,BAE Y S,et al.A direct role for dual oxidase in Drosophila gut immunity[J].Science,2005,310(5749):847-850.
[29] HA E M,OH C T,RYU JH,et al.An antioxidant system required for host protection against gut infection in Drosophila[J].Developmental Cell,2005,8(1):125-132.
[30] LIN P W,MYERS L E S,RAY L,et al.Lactobacillus rhamnosus blocks inflammatory signaling in vivo via reactive oxygen species generation[J].Free Radical Biology and Medicine,2009,47(8):1205-1211.
[31] JONES R M,MERCANTE JW,NEISH A S.Reactive oxygen production induced by the gut microbiota:pharmacotherapeutic implications[J].Current Medicinal Chemistry,2012,19(10):1519-1529.
[32] NEISH A S.Redox signaling mediated by the gut microbiota[J].Free Radical Research,2013,47(11):950-957.
[33] NEISH A S,JONES R M.Redox signaling mediates symbiosis between the gut microbiota and the intestine[J].Gut Microbes,2014,5(2):250-253.
