一种酿酒酵母表达CDNA文库分离纤维质降解酶系基因的方法
2012-11-02华承伟谢凤珍于江傲
华承伟,谢凤珍,于江傲
(1.河南科技学院生命科技学院,河南新乡453003;2.河南科技学院新科学院,河南新乡453003)
一种酿酒酵母表达CDNA文库分离纤维质降解酶系基因的方法
华承伟1,谢凤珍2,于江傲1
(1.河南科技学院生命科技学院,河南新乡453003;2.河南科技学院新科学院,河南新乡453003)
目的:建立一种筛选自然界产纤维质降解酶系基因的新方法。方法:对酿酒酵母表达载体pYES2多克隆位点加入真核生物稀有限制性酶切位点SfiI进行改造,利用SMART技术,以大肠杆菌文库为转导,构建了拟青霉(Paecilomyces sp.H28)的酿酒酵母全长cDNA表达文库。结果:利用纤维质-刚果红染色法从文库筛选到多种纤维素和半纤维素降解酶系基因。结论:成功构建了拟青霉的酿酒酵母表达cDNA文库,加快了纤维质降解酶系基因的快速分离,也为其它相关基因的快速分离提供了有益的借鉴。
酿酒酵母,表达cDNA文库,纤维质降解酶,分离,拟青霉
1 材料与方法
1.1 材料与仪器
拟青霉(Paecilomyces sp.H28) 由河南科技学院分离工程实验室筛选并保藏;酿酒酵母INVSc1(Saccharomyces cerevisiae)及载体pYES2 Invitrogen;Escherichia coli strain JM109 Stratagene;无氨基酸酵母氮源(Yeast Nitrogen Base without Amino Acids,YNB) Difco;酵母提取物、胰蛋白胨 Oxoid;桦木木聚糖 Sigma;TRIzol ReagentInvitrogen;Oligotex mRNA Mini Kit Qiagen;PrimeScript Reverse Transcriptase(RNase H-),Advantage 2 PCR KitTaKaRa;T4 DNA连接酶及其它限制性内切酶 NEB;质粒提取试剂盒 北京天根生化科技;其它试剂 均为分析纯。
1.2 pYES2载体的改造
1.2.1 寡核苷酸单链合成 根据pYES2载体酶切位点,分别合成内部具有EcoRI酶切位点且两端带有HindIII和XhoI两个酶切位点粘末端及两端分别引入SfiI A及SfiI B酶切位点的末端磷酸化的寡核苷酸链。
1.2.2 双链DNA合成 用STE缓冲液溶解两条寡核苷酸链,浓度为100μmol/L,分别取等体积的两链溶液50μL混合,置于PCR仪上95℃加热10min,然后缓慢冷却到室温,产物于4℃暂时贮存或-70℃保存。
1.2.3 连接及转化 取约100ng经HindIII和XhoI双酶切的pYES2载体和双链连接产物1.2pmol,T4DNA连接酶1μL,10×连接酶缓冲液1μL,加超纯水至总体积10μL,于16℃连接2h。连接产物直接化学转化法转化JM109大肠杆菌感受态细胞,铺制LB/Amp平板后于37℃培养过夜(12~16h),挑取10个单克隆经培养后送交测序公司进行序列测定。测定序列正确的载体命名为pYES2G。
1.2.4 SfiI双酶切位点的改造 合成两端带有6个保护碱基的EcoRI酶切位点的引物,PCR扩增长度约300bp不具有EcoRI酶切位点的任意一基因片段,PCR产物经琼脂糖凝胶回收,EcoRI酶切后连接经同样酶切及去磷酸化的pYES2G载体,连接产物转化大肠杆菌后,经PCR验证及SfiI酶切验证后,正确的载体命名为pYES2-GSL。
1.3 酿酒酵母表达cDNA文库的构建
1.3.1 总RNA及mRNA的提取 拟青霉(Paecilomyces sp.H28)经平板活化后,接种液体培养基于50℃,180r/min条件下培养4d,培养基组成(g/L):蛋白胨10.0;酵母粉10.0;MgSO4·7H2O 0.2;K2HPO40.87;KH2PO40.68;桦木木聚糖20.0;大麦葡聚糖5.0;CMC-Na 5.0;pH6.5。提取RNA所用的器皿及材料按无RNA酶条件处理。抽滤并用冰冷的DEPC处理的水洗涤菌丝体,称取0.1~0.3g菌丝体于液氮中快速研磨至无颗粒感,中间不断加入液氮防止液氮挥发干净,研磨后的菌丝体快速移入Trizol中(每0.1g菌丝体加入1mL Trizol),以下操作按Trizol说明书进行。琼脂糖凝胶变性电泳检测总RNA质量,紫外检测RNA纯度与含量后,利用Oligotex mRNA Mini Kit提取mRNA,具体操作参照试剂盒操作说明。
1.3.2 第一链全长cDNA的合成 第一链全长cDNA合成所需寡核苷酸引物SMART IV Oligonucleotide和CDS III/3’PCR Primer参照Creator SMART cDNA Library Construction Kit(Clontech),取500ng mRNA,利用PrimeScript Reverse Transcriptase(RNase H-)合成第一链全长cDNA,具体操作见试剂盒说明书。
1.3.3 长距离PCR(LD PCR)扩增全长cDNA 双链全长cDNA合成所需引物5’PCR Primer和CDS III/3’PCR Primer参照Creator SMART cDNA Library Construction Kit(Clontech)。取第一链产物2μL,利用Advantage 2 PCR Kit(TaKaRa)合成双链全长cDNA,PCR产物经蛋白酶K消化及SfiI酶切后经CHROMA SPIN-400 Column分级分离和乙醇沉淀后溶于8μL去离子水,取1μL检测DNA浓度,产物立即用于下一步连接或贮存于-20℃备用,具体操作见试剂盒说明书。
1.3.4 双链cDNA与载体pYES2-GSL的连接 取cDNA 2μL,pYES2-GSL(0.1μg)1μL,10×连接酶缓冲液0.5μL,T4DNA连接酶(400U/μL)0.5μL,去离子水1μL,总体积5μL,16℃连接过夜。连接产物加入95μL DEPC处理的去离子水和1.5μL糖原(20μg/μL),280μL冰冷的95%乙醇,混匀后于-70℃沉淀2h,15000r/min离心20min,弃去上清,待残存乙醇挥发干净后,沉淀溶于5μL DEPC处理的去离子水。
1.3.5 重组质粒电转化E.coli JM109 电转化感受态细胞及转化操作参照文库构建试剂盒,取1μL转化产物,用LB液体培养基稀释至10-6,取100μL涂布LB/Amp平板,检测重组率及文库滴度。根据文库滴度,涂布LB/Amp平板进行文库扩增,每个平板加入2mL灭菌的去离子水,用涂布棒轻轻刮掉菌落后用于质粒的提取。同时做一个没有连接载体的空白对照。
1.3.6 重组质粒电转化酿酒酵母INVSc1 酿酒酵母电转化感受态细胞制备参照《精编分子生物学实验指南》[5],取稀释后的重组质粒5μL(20~50ng),加入冰上融化的80μL酿酒酵母感受态细胞,轻轻混匀后,冰上放置5min,转入冰冷的0.2cm的电击杯中进行电击,电转参数设置:C=25μF;PC=200ohm,V=1.5kV。电击后立即在电转杯中加入1mL冰冷的1mol/L山梨醇。取1μL电击产物,加入100μL 1mol/L山梨醇涂布含有2%棉籽糖的SC-U选择性平板上(pYES2,Catalog no.V825-20,Version J,2004),用于计算文库滴度;其余转化产物分别取200μL涂布相应的SC-U选择性平板,用于文库扩增。30℃培养2~3d,挑取10个菌落PCR检测重组率。每个平板加入5mL 15%的甘油,轻轻刮掉菌落,于-70℃贮存备用。
1.4 酿酒酵母表达文库的筛选
取适当稀释的酿酒酵母表达文库,涂布双层筛选平板(含有2%半乳糖,2%棉籽糖,1%CMC-Na或木聚糖或大麦葡聚糖的SC-U诱导选择性平板),30℃,培养3d。每个平板加入适量的0.1%刚果红溶液,缓慢振荡染色1h,再用1mol/L的NaCl溶液洗脱,每次洗脱时间为5min,洗脱两次,观察有无透明圈。
2 结果与分析
2.1 pYES2载体的改造
限制性内切酶SfiI在真核生物中极少见到,因此在对酿酒酵母表达载体pYES2多克隆位点(MCS)的改造中引入SfiI酶切位点,合成两条互补末端磷酸化的寡核苷酸链碱基组成见图1。经等摩尔混合,加热退火后,合成双链DNA,连接HindIII和XhoI双酶切的载体pYES2,经测序鉴定,连接正确的载体命名为pYES2G。

图1 两条互补寡核苷酸链碱基组成Fig.1 The base composition of two complementary oligonucleotide chain
由于SfiI限制性内切酶在酶切时,两个SfiI位点之间至少需要170bp左右的核酸片段才能获得最佳的酶切效率[6],所以合成两端带有EcoRI酶切位点的引物,PCR扩增约300bp的插入片段(stuffer fragment),酶切后连接pYES2G,经SfiI酶切鉴定后,正确的载体命名为pYES2-GSL(图2A,图2B)。
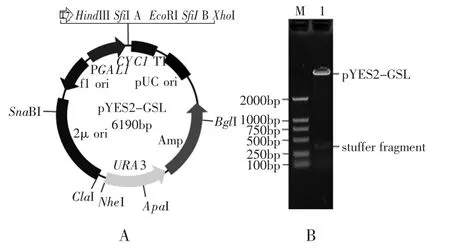
图2 载体pYES2-GSL图谱(A)和酶切分析(B)Fig.2 The features of the pYES2-GSL vecto(rA)and enzyme digestion analysis of recombinant pYES2-GSL(B)注:M:marker;1:pYES2-GSL digested with SfiI。
2.2 酿酒酵母表达cDNA文库的构建

图3 总RNA甲醛变性琼脂糖凝胶电泳分析(A)及双链cDNA电泳(B)Fig.3 Analysis of total RNA by formaldehyde-agarose gel electrophoresi(sA)and ds cDNA by agarose gel electrophoresi(sB)注:M:marker;A1:total RNA;B1:ds cDNA。
2.2.1 总RNA及mRNA的提取及双链cDNA合成 利用Trizol试剂成功从拟青霉(Paecilomyces sp.H28)提取得到总RNA,其1.5%琼脂糖凝胶变性电泳显示(图3A),28S约为18S的两倍。其OD260/OD280为1.90,OD260/OD230为1.94,说明提取的总RNA质量较好,利用Oligotex mRNA Mini Kit提取mRNA,立即用于反转录合成全长cDNA第一链,或分装后贮存于-70℃备用。取约500ng mRNA利用SMART(Switching Mechanism At 5’end of RNA Transcript)法合成全长cDNA第一链后,用Advantage 2 PCR Kit扩增形成双链cDNA,PCR产物主要集中在0.5~5kb之间(图3B),符合建库要求。
2.2.2 双链cDNA与载体pYES2-GSL的连接与转化 双链cDNA经蛋白酶K消化及SfiI酶切后,经CHROMA SPIN-400 Column分级分离后(图4),收集大于500bp的核酸片段,经乙醇沉淀浓缩后,连接载体pYES2-GSL。连接产物经电转化感受态细胞E.coli JM109,经文库滴度检验,未扩增文库滴度为2.2×108cfu/mL,重组率为98%,说明以大肠杆菌为宿主的cDNA文库具有很好的代表性。

图4 cDNA的分级分离Fig.4 cDNA size fractionation by CHROMA SPIN-400 Column
2.2.3 酿酒酵母表达cDNA文库的构建及筛选 扩增后的文库经质粒提取后,经适当稀释,分批电转化酿酒酵母,为使文库具有更好的代表性,需使酿酒酵母文库库容达106以上。本研究酿酒酵母电转化转化率达3.1×106/μg DNA,重组率达96%。文库经适当稀释后涂布纤维质SC-U诱导平板,经刚果红染色及脱色后,从文库中筛选到了纤维素酶、木聚糖酶和葡聚糖酶等多种纤维质降解酶系基因。酿酒酵母INSVc1本身所产胞外酶基本不含纤维质降解酶系,图4显示了筛选到的木聚糖酶菌株与阴性对照菌株的刚果红染色结果,虽然pYES2载体及重新改造的pYES2-GSL载体不具有分泌表达功能,但可以通过构建全长cDNA文库,借助蛋白本身信号肽进行分泌表达,虽然这种分泌表达能力较差,但借助一定的筛选手段并结合序列测定,可以检测到目标蛋白,从而筛选到有用的目标基因。

图5 重组酿酒酵母阴性菌株(A)和木聚糖酶重组酿酒酵母阳性菌株木聚糖刚果红平板(B)Fig.5 The analysis of recombinant S.erevisiae negative contro(lA)and xylose recombinant S.erevisiae by xylose-Congo red plate(B)
3 结论
酿酒酵母(S.erevisiae)在酿酒业和面包业的使用已有数千年的历史,被认为是GRAS(generally recognized as safe)生物[7],其完整的基因序列也于1996年完成测序,酿酒酵母具有大肠杆菌等原核生物繁殖快、易培养的优点,是最早应用于基因工程的酵母,是基因表达系统的优良宿主菌,至今已广泛用于表达各种外源基因[8-9]。pYES2为酵母-大肠杆菌多拷贝的穿梭质粒,是T7强启动子控制下的表达载体,通过SMART法合成全长cDNA,克隆进改造的pYES2-GSL载体,利用基因自身编码信号肽序列,可实现大部分基因尤其是丝状真菌基因的分泌表达,从而为基因文库的筛选提供简便的方法。整个技术的关键是初级大肠杆菌文库的滴度要达到108cfu/mL以上及酿酒酵母的转化率至少要达到105/μg DNA,以防文库信息的丢失。本研究利用构建的拟青霉(Paecilomyces sp.H28)酿酒酵母表达文库筛选得到木聚糖酶、葡聚糖酶和纤维素酶等多种纤维质降解酶系相关基因,同时,酿酒酵母不产生胞外葡糖糖苷酶,也可利用4-甲基伞形基-β-D-葡萄糖苷(4-MUG)等底物显色法筛选葡萄糖苷酶等糖苷水解酶类。同样,可利用三嗪染料活性蓝F3GA(Cibacron Blue F3GA)染料标记木聚糖、葡聚糖和淀粉[10],雷马亮蓝R(RBB)、活性艳红等标记木聚糖、菊粉和淀粉[11-12]等作为底物来筛选其它多糖降解酶类的基因。
[1]阎伯旭,高培基.纤维素酶分子结构与功能研究进展[J].生命科学,1995,7(5):22-25.
[2]Yoon J J,Kim K Y,Cha C J,et al.Purification and characterization of thermostable beta-glucosidase from the brownrot basidiomycete Fomitopsis palustris grown on microcrystalline cellulose[J].J Microbiol,2008,46(1):51-55.
[3]Mohamed G,Issam S.Fungus β-glycosidases:immobilization and use in alkyl β-glycoside synthesis[J].J Mol Catal B-Enzym,2004,299(1-6):89-94.
[4]Schwarz W H.The cellulosome and cellulose degradation by anaerobic bacteria[J].Appl Microbiol Biotechnol,2001,56(5-6):634-649.
[5]Ausubel M M,Brent R,Kingston R E,et al.Short protocols in molecular biology(5th Edition)[M].New York:Greene Pub Associates,2002:512-514.
[6]Wentzella L M,Halford S E.DNA looping by the SfiI restriction endonuclease[J].J Mol Biol,1998,281(3):433-444.
[7]Hitzeman R A,Leung D W,Perry L J,et al.Secretion of human interferons by yeast[J].Science,1983,219(4585):620-625.
[8]Romanos M A,Scorer C A,Clare J J.Foreign gene expression in yeast:A review[J].Yeast,1992,8(6):423-488.
[9]Ye N B,Xu H,Wang Y,et al.Cloning and characterization of a novel Δ12-fatty acid desaturase gene from the tree Sapium sebiferum[J].Biotechnol Lett,2007,29(6):959-964.
[10]Ten L N,IM W-T,Kim M-K,et al.Development of a plate technique for screening of polysaccharide-degrading microorganisms by using a mixture of insoluble chromogenic substrates[J].J Microbiol Meth,2004,56(3):375-382.
[11]Castro G R,Baigorí M D,Sineriz F.A plate technique for screening of inulin degrading microorganisms[J].J Microbiol Meth,1995,22(1):51-56.
[12]Ten L N,Im W-T,Aslam Z,et al.Novel insoluble dye-labeled substrates for screening inulin-degrading microorganisms[J].J Microbiol Meth,2007,69(2):353-357.
Method of isolating fiber degrading enzymes gene by S.cerevisiae expression cDNA library
HUA Cheng-wei1,XIE Feng-zhen2,YU Jiang-ao1
(1.School of Life Science and Technology,Henan Institute of Science and Technology,Xinxiang 453003,China;2.College of Xinke,Henan Institute of Science and Technology,Xinxiang 453003,China)
Object:Establish a method of isolating fiber degrading enzymes gene.Methods:pYES2 expression vector of Saccharomyces cerevisiae was reconstructed by adding the rare SfiI restriction site in eukaryotes into multiple cloning site(MCS).Using SMART technology,through Escherichia coli cDNA library,constructed the Saccharomyces cerevisiae full-length cDNA express library of Paecilomyces sp.H28.Results:Using the method of fiber-Congo red stain,screened a variety of cellulose and hemicellulose degrading enzymes genes from the library.Coclusion:Constructed S.cerevisiae cDNA expression library of Paecilomyces successfully. The method speeded up the rate of isolation of fiber-degrading enzymes genes,and provided the beneficial reference for other related genes isolation.
Saccharomyces cerevisiae;expression cDNA library;fiber-degrading enzymes;isolation;Paecilomyces sp.
Q789
A
1002-0306(2012)03-0167-04
纤维质是地球上最丰富的可再生的生物质资源,纤维质主要包括纤维素和半纤维素,对其的综合利用在解决能源危机、粮食短缺和环境污染等重大问题上具有深远的意义。但由于纤维质底物的高度复杂性等原因,纤维质的生物转化、利用与实际应用一直有相当的距离,纤维质的综合利用是世界难题,也是当前高技术领域竞争的焦点。纤维质很难被酶解,其酶解的转换率数比淀粉低1~2个量级[1]。因此怎样提高其酶解效率就成为本领域中的关键,微生物来源尤其是真菌来源的纤维素和半纤维素酶系在纤维质的生物降解中具有重要的作用,在食品、饲料、化工、制药及制浆造纸等工业领域中具有非常广阔的应用前景[2-4],近年来已受到研究者的广泛关注,但传统的菌种选育和酶学研究在这方面成就不大。当今分子生物学的快速发展及生物工程的兴起,克隆及表达高效、性能优良的纤维质降解酶系及利用基因工程和蛋白质工程重组、改造纤维质降解酶系等,为纤维质降解酶系注入了新的活力。迄今,已有上百种相关基因得到克隆和表达,真菌基因往往由于内含子的存在,导致使用传统的克隆方法耗时、费力且速度较慢。近年来,随cDNA文库构建技术的成熟,从cDNA文库出发,大大加快了相关基因的筛选和克隆,但一般以细菌和噬菌体构建的文库存在筛选方法复杂及工作量大等缺点,以酵母等真菌直接构建的文库也存在转化效率低,库容量小等缺点,在实际应用中也存在较大问题。本研究利用重新改造的酿酒酵母(Saccharomyces cerevisiae)穿梭表达载体pYES2构建拟青霉(Paecilomyces sp.H28)表达cDNA文库,利用透明圈法成功筛选到木聚糖酶、葡聚糖酶和纤维素酶等多种纤维质降解酶系基因。
2011-03-03
华承伟(1972-),男,博士,副教授,研究方向:食品生物技术。
河南科技学院重点科研项目资助基金。
