解淀粉芽孢杆菌TF28抗菌粗蛋白的初步研究1)
2011-08-07孟利强李晶张淑梅赵晓宇王玉霞曹旭
孟利强,李晶,张淑梅,赵晓宇,王玉霞,曹旭
(黑龙江省科学院 微生物研究所,哈尔滨 150010)
随着对植物病害防治要求的提高,传统抗生素作为杀菌剂显现许多弊端,如容易产生耐受性或抗性菌株,有毒性等,现在人们越来越多地把注意力投入到一类新型物质上,即抗菌蛋白或抗菌肽[1-3]。抗菌蛋白或抗菌肽是许多生防微生物都能分泌的一类活性物质,它具有抑制病菌的广谱性和高效性、对人畜无毒、对环境无污染的优良特性,且病菌对其不易产生抗性[4]。因此,抗菌蛋白在新型生物农药的开发中具有较好的利用价值。
解淀粉芽孢杆菌(Bacillus amyloliquefaciens)是一类可产生丰富胞外拮抗物质的革兰氏阳性细菌,从中分离出具有较好生防应用价值的拮抗物质,并对其进行开发利用正成为解淀粉芽孢杆菌应用研究的热点[5-6]。
解淀粉芽孢杆菌TF28是从大豆根部分离出的一株拮抗内生细菌,经测定,该菌产生的抗菌蛋白具有广谱高效的抑菌活性。本文研究对其抗菌粗蛋白的理化性质和抑菌作用进行了研究,以期为该蛋白的进一步开发利用提供科学依据。研究生防细菌抗菌蛋白的性质及其对植物病原菌的拮抗作用,一方面能充分发掘有益微生物资源,将其用于植物病害生物防治;另一方面为研究抗菌蛋白的分子遗传基础、克隆抗菌蛋白基因以及构建转基因工程菌奠定理论基础[7-8]。
1 材料与方法
1.1 实验材料
拮抗菌株解淀粉芽孢杆菌TF28为本研究室分离保存,12株供试植物病原菌为本研究室保存。
1.2 实验仪器
Sigma低温台式离心机、UV1102紫外分光光度计、恒温培养箱、超净工作台、涡旋振荡器、HZQ-QX全温振荡器、HWS24电热恒温水浴锅、OLYMPUS万用显微镜。
1.3 实验方法
1.3.1 TF28菌株生长曲线与抑菌时程曲线
将活化好的TF28菌种按5%接种量接入50mL NYD培养液中,在30℃,转速为180rpm摇床上振荡培养,每隔8h取2mL培养液两份,一份用紫外分光光度计测定OD600吸收值,另一份经6000rpm离心10min,收集上清液,采用杯碟法[9]测定抑菌活性。以培养时间为横坐标,发酵液的OD600吸收值和抑菌活性为纵坐标作图,分析TF28菌株培养时间与抗菌物质抑菌活性的关系。
1.3.2 抗菌粗蛋白的制备
取1000mL TF28菌株发酵上清液,缓慢地加入固体硫酸铵至20%饱和度,4℃静置2h,6000rpm离心30min,分别收集上清和沉淀。在收集的上清液中再次加入固体硫酸铵,同法调节至40%,60%,80%,100%饱和度。收集每个饱和度下的沉淀物,分别将其溶解在0.02mol/L磷酸缓冲液(pH6.8)中,经透析及聚乙二醇包埋浓缩后定容至10mL,测定各组分的抗菌活性,活性强的组分即为制备的抗菌粗蛋白,用于以下研究。
1.3.3 抗菌粗蛋白性质研究
热稳定性:将抗菌粗蛋白分别置于50℃、70℃、90℃和100℃水浴中处理30min,各取100μL处理后样品用杯碟法测定其抑菌活性,以未经热处理的抗菌粗蛋白为对照,将对照的抑菌活性定为100%(下同)。
pH稳定性:将抗菌粗蛋白用1mol/L HCl溶液和1mol/L NaOH 溶液分别调节pH 值至1.0、3.0、5.0、7.0、9.0、11.0、13.0,置4℃过夜保存,再将各pH 值调回至中性后测定其抑菌活性。
紫外线稳定性:将抗菌粗蛋白置于28WUV灯下,距离10cm处分别照射l、2、3、4、5h,测定其抑菌活性。
蛋白酶稳定性:将抗菌粗蛋白分别用胰蛋白酶、蛋白酶K在37℃下处理30min,酶解反应浓度为1mg/mL,测定其抑菌活性。
1.3.4 抗菌粗蛋白的抑菌作用
抗菌粗蛋白对水稻恶苗病菌孢子萌发的影响:制备不同浓度的抗菌粗蛋白,取抗菌粗蛋白、病原菌孢子(106个/mL)及PD液体培养基各300μL于1.5mL的EP管中,抗菌粗蛋白终浓度分别为553.28、276.64、138.32、69.16、34.58、17.29μg/mL。25℃静置培养12h,光学显微镜观察,统计视野内孢子的总数和萌发的孢子数。对照采用75%百菌清可湿性粉剂800倍稀释液。每个处理做3个重复,计算孢子萌发率(孢子萌发率=萌发孢子数/孢子总数)×100%。
抗菌粗蛋白对水稻恶苗病菌菌丝形态的影响:在PD培养基平板中心接入病原菌块,采用滤纸片法[10]测定抗菌粗蛋白对菌丝形态的影响:每个滤纸片上加10μL的抗菌粗蛋白,对照加无菌水。培养24h,用显微镜观察抑菌圈周围菌丝形态的变化。
抗菌粗蛋白对病原菌菌丝生长量的影响:PD液体培养基(50/250mL)中接入病原菌孢子液至终浓度104个/mL,于28℃,150rpm摇床培养,培养5h后加入抗菌粗蛋白 (终浓度为138.32μg/mL),继续培养7d后,收集菌丝,烘干称重,计算菌丝生长量。
2 结果与分析
2.1 TF28菌株生长曲线与抑菌时程曲线
在TF28菌株液体发酵培养过程中每隔8h取样一次,分别测定培养液OD600吸收值及上清液抑菌活性。以培养时间为横坐标,OD600吸收值和抑菌活性为纵坐标作图,分析TF28的培养时间与抗菌物质抑菌活性的关系。结果如图1所示,在初始的24h内菌密度呈对数增长,随后进入稳定期。其发酵上清液抑菌活性随着菌密度的增加而增大,但其活性在48h时达到最大值,相对于菌密度的增加,抑菌物质的产生存在滞后性,48h过后其抑菌活性稍有下降,可见TF28菌株培养至48h时抑菌活性最强。
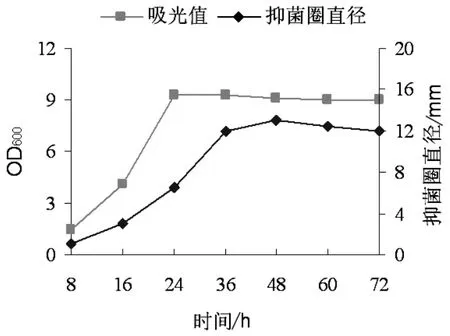
图1 菌株TF28发酵液OD600值与抑菌活性曲线
2.2 抗菌粗蛋白的制备
抗菌粗蛋白的获得采用硫酸铵盐析法[11],硫铵饱和度为20%时,其粗提物抑菌活性最弱;(图2)当酸铵盐析饱和度为40%时,粗提物抑菌活性最强;而当硫酸铵盐析饱和度为60%,80%和100%时,粗提物的抑菌活性都远低于40%饱和度,由此确定,沉淀抗菌粗蛋白的最佳硫酸铵饱和度为20%~40%。

图2 不同饱和度的硫酸铵盐析对抗菌粗蛋白抑菌活性的影响
2.3 抗菌粗蛋白性质研究
热稳定性:该抗菌粗蛋白经50℃和70℃处理30min活性几乎不变(表1),而经90℃和100℃处理30min活性有所下降,抑菌率分别为原有活性的93.3%和85.9%,由此可见该抗菌粗蛋白具有一定的热稳定性。
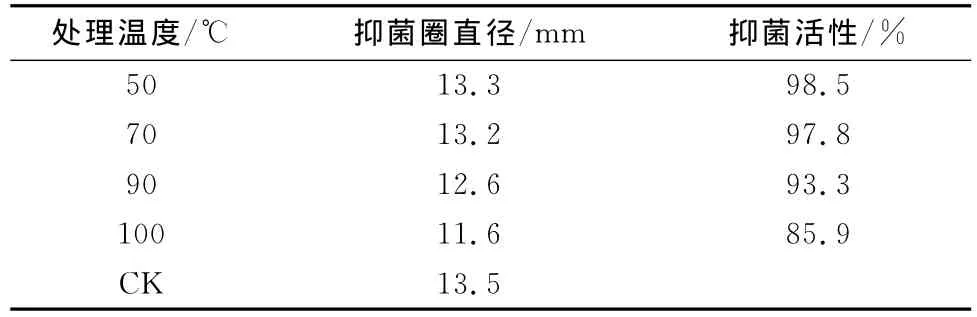
表1 抗菌粗蛋白的热稳定性
pH的稳定性:该抗菌粗蛋白在较广pH(3~11)范围内比较稳定,活性均能保持在95%以上(表2),仅在强酸(pH1)和强碱(pH13)的环境中活性有所下降,分别为原有活性的86.9%和82.5%。

表2 抗菌粗蛋白的pH稳定性
对紫外线的稳定性:该抗菌粗蛋白经紫外线照射后不会失活,对紫外线不敏感。
对蛋白酶的稳定性:该抗菌粗蛋白经胰蛋白酶、蛋白酶K分别在37℃下处理30min,结果显示该抗菌粗蛋白对胰蛋白酶不敏感;而蛋白酶K处理使其活性降低,但仍能保持原有活性的79.3%,即对蛋白酶K具有一定的耐受性。
2.4 抗菌粗蛋白的抑菌作用
2.4.1 抗菌粗蛋白对水稻恶苗病菌孢子萌发的影响
用不同浓度抗菌粗蛋白处理水稻恶苗病菌孢子,发现病菌孢子的萌发率随抗菌粗蛋白浓度的改变而改变,浓度越高孢子的萌发率越低,结果见图3。当抗菌粗蛋白的浓度为17.29μg/mL时,孢子的萌发率为99.46%;而当抗菌粗蛋白的浓度为553.25μg/mL时,病菌孢子的萌发率仅为20.1%。该抗菌粗蛋白抑制水稻恶苗病菌孢子萌发的有效中浓度(EC50)为138.32μg/mL。

图3 不同浓度抗菌粗蛋白病原孢子萌发的影响
2.4.2 抗菌粗蛋白对水稻恶苗病菌菌丝形态的影响
在PDA平板上,经抗菌粗蛋白处理的水稻恶苗病菌菌丝变形以至破裂。显微观察发现,未经处理的菌丝长且致密,粗细均一,菌丝内部物质分布均匀,折光一致(图4A)。而经过抗菌粗蛋白处理12h后,病原菌菌丝多处出现膨大,菌丝粗细不均;在处理24h后菌丝出现肿胀,并且菌丝末端出现泡囊结构。随着处理时间的延长,菌丝的畸形程度加剧(图4B)。

图4 抗菌粗蛋白对水稻恶苗病病原菌菌丝形态的影响
2.4.3 抗菌粗蛋白对不同病原菌菌丝生长量的影响
将终浓度138.32μg/mL的抗菌粗蛋白分别接入到12株活化好的病原菌PD液体培养基中,28℃培养7d,结果抗菌粗蛋白对12株病原菌菌丝生长均有抑制作用,对照组菌丝大量生长,实验组仅见少量菌丝。其中对大豆菌核病菌(Sclerotinia sclerotiorum)抑制率最低为71.3%,对大豆根腐病菌(Fusarium oxysporumf.sp.vedolens)和水稻恶苗病菌(Fusarium moniliforme)抑制率较高分别为93.2%和94.3%(表3)。

表3 抗菌粗蛋白对病菌菌丝生长量的抑制率
3 结论
生防解淀粉芽孢杆菌TF28是从大豆根部分离出的一株内生细菌,具有良好的实验室生测活性,能对多种植物病原菌的生长起到抑制作用。从TF28菌株生长曲线与抑菌时程曲线看,菌体在24h达到稳定期,其发酵上清液抑菌活性在培养48h后达到峰值且抑菌活性没有随菌体生长衰退而明显下降,说明抗菌活性物质具有较高的稳定性。利用20%~40%饱和度的硫酸铵从其发酵上清液中盐析可获得抗菌粗蛋白,该粗蛋白具有一定的热稳定性,100℃处理30min后其抑菌活性仍为原有抑菌活性的85.9%;且在较广pH(3~11)范围内的比较稳定,活性均能保持在95%以上;对紫外线、胰蛋白酶不敏感,对蛋白酶K具有一定的耐受性;该抗菌粗蛋白对已测定的12种真菌均有抑制作用,对大豆根腐病菌和水稻恶苗病菌的抑制作用尤为明显。其对水稻恶苗病菌孢子萌发的有效抑制中浓度(EC50)为138.32μg/mL。此外,该抗菌粗蛋白能破坏病原菌菌丝结构,导致其畸形,如菌丝肿胀和末端出现泡囊结构等。
综上所述,TF28菌株所产抗菌蛋白具有较高的理化稳定性及其广谱高效的抑真菌活性,具有良好的应用前景。对其抗菌蛋白有待于进一步开展分离纯化研究,从而进行抗菌蛋白拮抗基因的克隆及其表达调控研究,为高效、多功能生防基因工程菌的构建和抗病转基因植物的获得提供技术支撑[12-13]。
[1]程亮,游春平,肖爱萍等.拮抗细菌的研究进展[J].江西农业大学学报,2003,25(5):732-737.
[2]齐东梅,梁启美,惠明,等.棉花枯萎、黄萎病拮抗芽孢杆菌的抗菌蛋白特性[J].微生物学通报,2005,32(4):42-46.
[3]宋永燕,李平,郑爱萍,等.生防细菌LM-3的鉴定及其抗菌蛋白的研究[J].四川大学学报(自然科学版),2006,43(5):1110-1115.
[4]张少飞,李敏,邢志国,等.产抑菌物质SDLH菌株的鉴定及发酵产物稳定性研究[J].生物技术,2009,19(5):56-61.
[5]车晓曦,李社增,李校堃,等.1株解淀粉芽孢杆菌发酵培养基的设计及发酵条件的优化[J].安徽农业科学,2010,38(18):9402-9405.
[6]郝建安,曹志辉,赵凤梅,等.解淀粉芽孢杆菌NK10.BAhjaWT抑真菌作用的研究[J].微生物学通报,2008,35(6):903-908.
[7]Sutyak K E;Wirawan R E;Aroutchva A A,Isolation of the Bacillus subtilis antimicrobial peptide subtilosin from the dairy product-derived Bacillus amyloliquefaciens 2008,106(06):1067-1074.
[8]Wu W S;Wu H C;Li Y L Potential of Bacillus amyloliquefaciens for control of Alternaria cosmosa and A.patula of Cosmos sulfurous(Yellow Cosmos)and Tagetes patula(French Marigold)[J].Journal of Phytopathology,2007,12:670-675.
[9]纪兆林,唐丽娟,张清霞,等.地衣芽孢杆菌 W10抗菌蛋白的分离纯化及其理化性质研究[J].植物病理学报,2007,37(3):260-264.
[10]梁启美,齐东梅,贾洁,等.棉花黄、枯萎病拮抗菌的筛选及抗菌蛋白B110-a的初步测定[J].植物保护,2005,31(5):35-39.
[11]李晶,杨谦,赵丽华,等.生防枯草芽孢杆菌B29菌株抗菌物质的初步研究[J].中国生物工程杂志,2008,28(2):59-65.
[12]Shiou-Huei Chao;Tzu-Hao Cheng Characterization of a Novel PepF-Like Oligopeptidase Secreted by Bacillus amyloliquefaciens 23-7A[J].Appl Envir Microbiol,2006.72(1):968-971.
[13]林东,徐庆,刘忆舟等.枯草芽孢杆菌SO113分泌蛋白的抑菌作用及抗菌蛋白的分离纯化[J].农业生物技术学报,2001,9(1):77-80.
