咖啡因与味觉感知
2011-02-08刘斯斯陈梦玲张根华
刘斯斯,陈梦玲,张根华
(1.苏州大学 基础医学与生物科学学院,江苏 苏州 215006;
2.常熟理工学院 生物与食品工程学院,江苏 常熟 215500)
咖啡因广泛存在于日常生活消费品中,例如咖啡,巧克力,可乐及各种茶叶等.咖啡因在人体和哺乳动物中能产生多种生理作用,但是关于咖啡因是如何引起苦味感知,对于味觉偏好是否有影响,味觉信号是如何传导等问题尚未完全阐明.本文根据已有报道,从咖啡因的生理作用、苦味感知及信号传导三个方面进行概述,以期为今后咖啡因的味觉感知相关研究提供线索.
1 咖啡因的生理作用
咖啡因是从茶叶、咖啡果中提炼出来的一种生物碱,含有甲基黄嘌呤类成分.它具有兴奋中枢神经系统、促进新陈代谢的作用,可提高血浆游离脂肪酸的浓度,促进脂肪分解[2].已有文献报道,长期饮用含咖啡因的食物会明显降低人类和哺乳动物的体重[2,3].
咖啡因对机体有着广泛的生理作用:首先,它是中枢神经系统的兴奋剂,适度使用能祛除疲劳和兴奋神经,临床上用于治疗神经衰弱和昏迷复苏.但是,大剂量或长期使用会对人体造成损害,特别是它具有一定的成瘾性,一旦停用会出现精神萎顿、浑身困乏疲软等各种戒断症状.此外,有研究发现,经常摄入咖啡因能有效预防帕金森病,其原因可能是咖啡因对脑基底核具有保护作用[4].其次,咖啡因对心血管系统、消化系统、内分泌系统以及骨骼肌与运动能力都产生影响.咖啡因能促进儿茶酚胺的合成和释放,所以被认为是具有正性作用的心血管药物,即使一次服用小剂量也能引起心率增快、血压升高[1].另外有研究表明,长期饮用咖啡因能显著降低2型糖尿病的发病率[5].在消化系统方面,咖啡因可刺激迷走神经胃支,促进胃酸分泌[6].在骨骼肌和运动能力方面,咖啡因能增加肌浆网细胞中钙离子浓度,因而增加肌纤维的伸缩性,提高运动能力.
目前已知咖啡因在细胞水平上有以下几种作用机制[7]:①是中枢腺苷受体(A)的拮抗剂;②能抑制磷酸二酯酶,使细胞内环核苷酸浓度升高,引起一系列后续反应;③能显著提高细胞内钙离子浓度;④对多种电压门控离子通道有作用,能显著抑制钾离子通道,对钠离子通道有较弱抑制作用,但对氯离子通道没有作用.
2 咖啡因与苦味感知
味觉感知能引导有机体识别和摄入各种营养物质,同时也能帮助机体避免摄入有毒物质.目前已知人类能识别5种基本味觉,即酸、甜、苦、咸、鲜.最近几年的研究表明人类还能识别其他的味觉,例如脂肪味、金属味、辣味等[8].其中,苦味的识别作为一种防御机制,能帮助动物避免摄入有毒物质,它在动物的长期进化过程中起着至关重要的作用.对于现代人类而言,苦味感知作为一种防御机制所起的作用越来越小,而个体对苦味敏感性的差异可能会影响饮食行为,进而影响健康.
咖啡因已被证明能引起人类和哺乳动物的苦味感知.Keast等[9]利用三点实验法(triangle test)测得人类对咖啡因的感知阈值(detection threshold)是(1.2±0.12)mM,另外还测得对其他两种苦味物质QHCl(盐酸奎宁)和PROP(6-N-丙基-2-硫尿嘧啶)的感知阈值分别为(0.0083±0.001)mM和(0.088±0.07)mM.有文献报道,对咖啡因的味觉感知能力还与测试者的性别相关.Hirokawa等[10]对日本108名男女大学生进行了不同浓度咖啡因测试,发现男性和女性的感知阈值不同.相对男性而言,女性具有更低的感知阈值,也就是说女性对于咖啡因引起的苦味具有更高的敏感性,这可能与位于舌前部的味觉受体密度相关.有文献表明,女性的菌状乳头和菌状味蕾都比男性要多[10].Frank等[11]通过双瓶喜好实验(Two bottle preference test)对黄金仓鼠进行了一系列不同浓度的咖啡因苦味刺激测试.结果显示,当咖啡因浓度在(3-10)mM变化时,厌恶程度急剧增加(见图1),由此认为黄金仓鼠对咖啡因的厌恶阈值是3mM.对大鼠进行的双瓶喜好实验显示,大鼠对咖啡因的厌恶阈值为10mM[7].另外,Laska等[12]通过对三种非人类的灵长类品系(松鼠猴,蜘蛛猴,豚尾猴)的研究,发现不同品系的动物对咖啡因的厌恶阈值也不相同.
对于苦味的感知是长期自然选择进化的结果,而进化的过程涉及遗传和变异两个方面.美国科学家Hansen等[13]研究了遗传力和基因的协同变异对苦味物质敏感性的影响.他们的研究对象包括在遗传上具有不同研究意义的同卵双胞胎和异卵双胞胎;检测的苦味物质包括PROP、SOA、奎宁盐酸和咖啡因.实验表明,对这些苦味物质的敏感性受到遗传和基因变异的双重影响,并且对不同苦味物质的感知是受不同基因调控的.

图1 对咖啡因、SOA、MgSO4、denatonium benzoate和盐酸奎宁的双瓶喜好实验.圆圈标注的点是各物质的厌恶阈值[11]
3 咖啡因的味觉信号转导
关于味觉产生的生理学基础目前已相对较清楚.味觉感知的产生,是由于食物中特定的味质分子和味细胞中的味受体结合.味细胞是具有类神经元特性的表皮细胞,能够去极化,释放神经递质,形成和传入神经相连的突触.味细胞存在于味蕾中,味蕾呈洋葱状,大约50-150个味细胞构成一个味蕾.味蕾存在于舌表面特定的乳头中,根据形态可分为菌状乳头、叶状乳头和轮廓状乳头.除舌表皮外,在软腭、咽、喉、食管和会厌中也存在味蕾.尽管各种味质分子都能被舌识别,但舌面上不同区域对于特定味质分子的敏感性不同[14].菌状乳头大部分位于舌前部,对于甜味和咸味较敏感.叶状乳头主要位于舌后侧边缘,对于苦味和酸味敏感.轮廓状乳头主要位于舌后部,主要感知苦味和鲜味.当味质分子和相应受体结合后,味细胞发生去极化引发动作电位,导致神经递质直接或间接传递到传入神经,最终将信号传入大脑引起感知[8].
常见苦味物质的信号转导,例如的士宁、放线菌酮、PROP和PTC都是通过与膜上的苦味受体(taste receptor family 2 member,T2Rs)相结合,引起一系列的级联反应,从而形成苦味感知.苦味受体T2Rs属于G蛋白偶联受体超家族,具有7次跨膜螺旋结构,含有3个胞内环和3个胞外环.当苦味物质与相应的受体结合后,会引发由味导素介导的两条信号转导通路:第一条信号通路是α-味导素→磷酸二脂酶(PDE)→cNMP.当苦味物质与T2Rs结合后,激活α-味导素,进而活化PDE,降低细胞内cNMP的浓度,从而调控cNMPs门控的离子通道活性.第二条通路是β-γ-味导素→PLC→IP3/DAG.IP3能在胞内与其受体结合,动员胞内钙库释放Ca2+,提高胞内 Ca2+浓度,从而引起苦味受体细胞的去极化[15].
关于咖啡因的味觉信号转导途径,目前还没有明确的定论,而对于它的研究却从未中断.由于咖啡因具有很强的脂溶性,能透过细胞膜,所以有观点认为咖啡因的味觉信号转导可能不依赖苦味受体T2Rs及其偶联的 G蛋白.最早提出这种观点的是纽约大学的Rosenzweig等[16],他们通过quench-flow技术,检测了小鼠味觉组织中环鸟苷酸cGMP的产生和降解.一系列实验表明,咖啡因透膜后可以抑制细胞内的PDE,并使环化酶活化,导致细胞内的cGMP浓度增加,进而使细胞去极化.Rosenzweig等[16]还提到一个非常重要的非选择性阳离子通道—味觉特异性的环核苷酸门控通道(taste-specific cyclic nucleotide-gated channel,CNGgust),该通道已经从大鼠味觉组织中被克隆.由于咖啡因刺激后,能引起细胞内环核苷酸浓度增加,所以他们认为CNGgust可能介导阳离子内流引起细胞去极化或者释放神经递质(见图2).然而,在研究果蝇的味觉感知时,却发现了咖啡因的受体—Gr66a,这可能是迄今为止发现的第一个咖啡因受体.该受体在果蝇的味觉受体神经元树突处表达,对于果蝇感知咖啡因是必需的.行为学研究表明,缺失Gr66a受体的果蝇对咖啡因引起的厌恶反应消失,同时咖啡因和茶碱都不能使之产生动作电位[17].
通过膜片钳技术可以观察到,一定浓度的咖啡因刺激对离体味细胞的钾离子通道功能具有很强的抑制作用,对钠离子通道的抑制作用相对较弱,而对氯离子通道没有作用.同时,钙影像技术表明咖啡因能显著增加细胞内钙离子的浓度[10].Nagatomo等[18]通过钙影像和电生理研究证明,咖啡因能活化小鼠的瞬时受体电位A1(transient receptor potential A1,TRPA1),从而介导咖啡因的味觉感知,但对人类的TRPA1通道却有抑制作用,提示人类与小鼠对咖啡因的感知机制可能有差异.
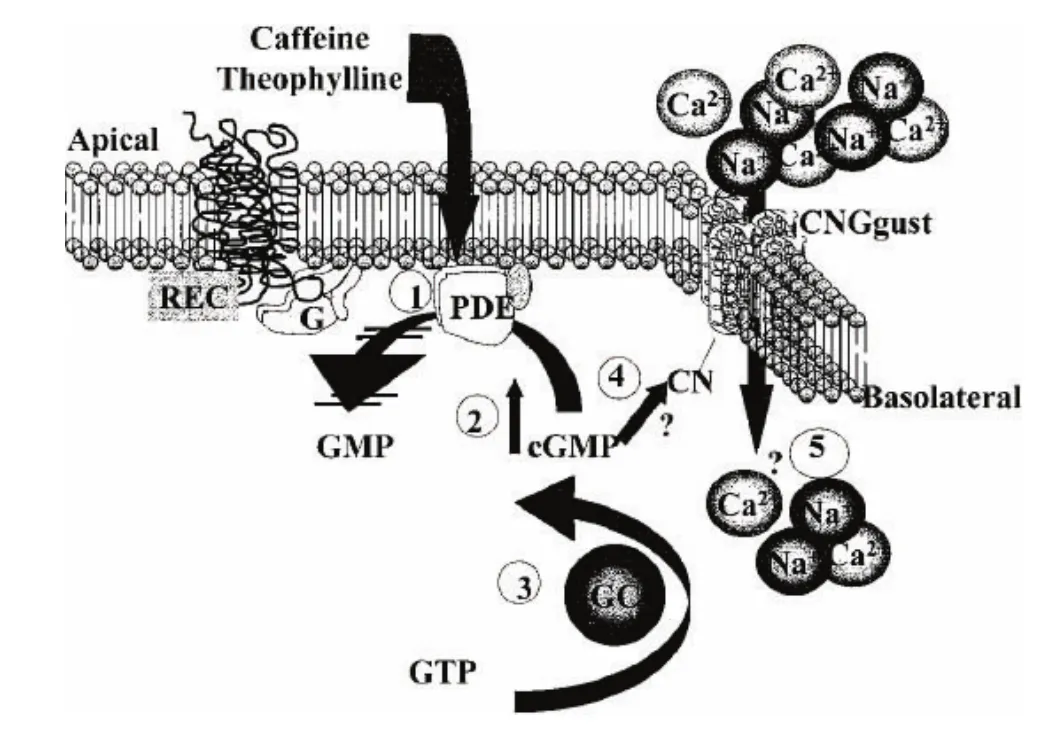
图2 咖啡因和茶碱的信号转导模式[16]
4 展望
咖啡因作为日常消费品中常见的物质,人们对其需求量已经越来越大.因此明确咖啡因的生理作用、咖啡因对于饮食偏好的影响以及咖啡因与味觉感知的生理学基础显得尤为重要.目前对咖啡因的研究手段主要包括行为学测试、电生理检测和钙影像技术等,研究还不够深入.另外,关于咖啡因是如何引起苦味并进行味觉信息传导的机制还没有定论.本文就目前相关的研究进行了综述,为进一步探讨咖啡因的功能和味觉信息传导机制奠定基础.
[1]易超然,卫中庆.咖啡因的药理作用和应用[J].医学研究生学报,2005,18:270-272.
[2]陆梦,张莉,丁雪瑞,等.咖啡因对小鼠空间学习记忆及体重的影响[J].西安交通大学学报(医学版),2010,31:648-650.
[3]Boozer CN.Herbal ephedra/caffeine for weight loss:a 6-month randomized safety and efficacy trial[J].Int J Obes Relat Metab Disord,2002,26:593-604.
[4]Ross GW,Abbott RD,Petrovitch H,et al.Association of coffee and caffeine intake with the risk of Parkinson disease[J].JAMA,2000,283:2674-2679.
[5]van Dam RM,Hu FB.Coffee consumption and risk of type 2 diabetes:a systematic review[J].JAMA,2005,294:97-104.
[6]Kwiecien S.Gastric analysis with fractional test meals(ethanol,caffeine,and peptone meal),augmented histamine or pentagastrin tests,and gastric Ph recording[J].J Physiol Pharmacol,2003,54:69-82.
[7]Zhao FL,Lu SG,Herness S.Dual actions of caffeine on voltage-dependent currents and intracellular calcium in taste receptor cells[J].Am J Physiol Regul Integr Comp Physiol,2002,283:R115-R129.
[8]Chaudhari N,Roper SD.The cell biology of taste[J].J Cell Biol,2010,190:285-296.
[9]Keast RSJ,Roper Jessica.A complex relationship among chemical concentration,detection threshold,and suprathreshold intensity of bitter compounds[J].Chem Senses,2007,32:245-253.
[10]Hirokawa K,Yamazawa K,Shimizu H.An Examination of Sex and Masculinity/Femininity as Related to the Taste Sensitivity of Japanese Students[J].Sex Roles,2006,55:429-433.
[11]Frank ME,Bouverat BP,MacKinnon BI,et al.The distinctiveness of ionic and nonionic bitter stimuli[J].Physiol Behav,2004,80.
[12]Laska M,Rivas Bautista RM,Hernandez Salazar LT.Gustatory responsiveness to six bitter tastants in three species of nonhuman primates[J].J Chem Ecol,2009,35:560-571.
[13]Hansen JL,Reed DR,Wright MJ,et al.Heritability and genetic covariation of sensitivity to PROP,SOA,quinine HCl,and caffeine[J].Chem Senses,2006,31:403-413.
[14]Sugimoto K.Introductory remarks on umami research:candidate receptors and signal transduction mechanisms on umami[J].Chem Senses,2005,30:i21-i22.
[15]胡玲玲,施鹏.苦味受体基因家族功能和演化研究的最新进展[J].科学通报,2009,54:2472-2482.
[16]Rosenzweig S,Yan wentao,Dasso M,et al.Possible novel mechanism for bitter taste mediated through cGMP[J].J Neurophysiol,1999,81:1661-1665.
[17]Moon SJ,Kottgen M,Jiao Y,et al.A taste receptor required for the caffeine response in vivo[J].Curr Biol,2006,16:1812-1817.
[18]Nagatomo K,Kubo Y.Caffeine activates mouse TRPA1 channels but suppresses human TRPA1 channels[J].Proc Natl Acad Sci U S A,2008,105:17373-17378.
