全公猪肉膻味的调控
2010-11-22柳东阳朱桂姿编译
柳东阳,朱桂姿 编译
(1.华中农业大学动物医学院,湖北 武汉 430070;2.河南省安阳县职业中专,河南 安阳 455112)
气味是肉品的重要感官特性,决定了消费者是否购买。猪肉气味很大程度上受到雄性膻味的影响,目前认为膻味主要由雄烯酮(androstenone)和粪臭素(skatole)构成。传统上对生产猪肉的小公猪进行阉割,目的在于增加胴体中的脂肪含量、减少攻击性和性行为、降低产生膻味的危险。现在消费者倾向于购买瘦肉,阉割得以保留原因主要是降低膻味和减少公猪攻击性。但阉割的优越性也因消费者对猪肉生产中动物福利的关注而受到质疑。此外,因改进了饲料转化率和提高了瘦肉率,饲养全公猪的效益更高。与阉猪相比,全公猪多聚不饱和脂肪酸和蛋白含量高,在营养上具有优势。
欧洲食品安全局(EFSA 2004)报告不同研究者对膻味的描述是:令人不愉快的气味、像尿味、兽味、汗味和粪味。欧洲六国一个研究小组发现:雄烯酮与尿味有关;粪臭素与粪臭味有关,还有少许萘的味道。即使通过大量测试和统计学对参与者的描述偏差做以纠正,膻味的感觉特性仍然非常复杂。原因可能是培训中缺乏对膻味有一个统一的定性和定量标准。依据个人敏感度,对于膻味的反应差异很大。猪肉膻味取决于膻味化合物合成和清除的速度。这些过程受到一系列相关代谢酶类表达的影响。
1 雄烯酮的合成与代谢
雄烯酮有较强的脂溶性,睾丸中合成后一部分进入血液,再进入脂肪,加热时产生膻味。一部分进入唾液,作为性外激素发挥作用。最后一部分进入肝脏经过一期和二期降解分泌入胆汁,经粪便排泄。
1.1 雄烯酮的合成
雄烯酮与其他睾丸同化激素一样产生于公猪睾丸间质细胞,此外肾上腺和卵巢中也可合成,因此母猪和阉猪血液中也可以检测到雄烯酮。细胞色素P450C17(CYPI7A1)和细胞色素b5(CYPB5)在合成雄烯酮时起着重要作用。雄烯酮来源于17-乙炔睾酮,经andien-β-合成酶合成。雄烯酮和其他睾丸激素的合成受控于下丘脑——垂体——性腺轴,在出生后2~4 周,下丘脑——垂体——性腺轴有一个短暂的活化,增加了循环中雄烯酮的含量。青年公猪雄烯酮合成较少,随着性成熟而逐渐增加。神经内分泌系统对青春期公猪睾丸间质细胞合成雄烯酮具有很大的决定作用。遗传背景对合成雄烯酮的强度也有很大影响,Bonneau 等于1987 年鉴定出了血浆中睾酮和黄体生成素(LH)含量正常,而脂肪中雄烯酮含量较低的成年公猪。
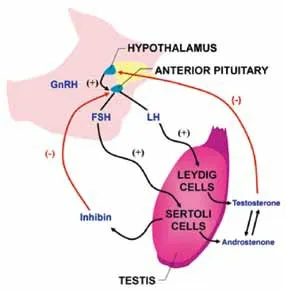
图1 雄烯酮合成调节示意图
1.2 雄烯酮的代谢
雄烯酮代谢主要在睾丸和肝脏,由3α-羟化类固醇脱氢酶、3β-羟化类固醇脱氢酶和二期结合酶(羟化类固醇磺基转移酶:SULT2A1 或 SULT2B1 和UDP-葡萄糖苷转移酶UGTs)参与。
在睾丸中合成的雄烯酮立即由羟化类固醇磺基转移酶(SULT2A1/SULT2B1)连上磺基。体内和体外试验表明,雄烯酮的代谢物为3α-雄烷醇和3β-雄烷醇,最后雄烷醇代谢为极性更强的结合型类固醇。
肝脏与睾丸中雄烯酮代谢不同的是,产生的以上2 种代谢物的百分比不同。雄烷醇经过二期代谢生成葡萄糖苷结合物和磺基结合物。羟化类固醇磺基转移酶是肝脏中16- 雄烯酮类固醇代谢的关键酶。最近发现,SULT2B1 也参与其中。此外,雄烯酮也可以通过SULT2B1 对第3 位的酮基烯醇化而加上一个磺基结合基团。Sinclair and Siquires(2005)等报道一些公猪的血浆中雄烯酮大部分是磺基化的,而Zamratskaia(2007)等报道称磺基化的雄烯酮要比游离的雄烯酮低很多。同样, Tuomola(1997)采用47%和100%的甲醇来分离结合和游离的雄烯酮,并未发现在血浆中有任何结合型的雄烯酮。今后应更进一步研究雄烯酮结合磺基之后对其消除和脂肪中积累的影响。
一部分雄烯酮可以经循环进入到颌下 腺。Both(1987)用H3标 记3α-雄烷醇,转化为雄烯酮,雄烯酮转化为3β-雄烷醇,雄烷醇保持不变。
2 粪臭素的合成与代谢
粪臭素有脂溶性和水溶性,在大肠合成吸收进入血液,一部分转入肝脏经一期和二期代谢,经胆汁排泄。另一部分进入脂肪产生膻味。
2.1 粪臭素生物合成
粪臭素来源于L-色氨酸,在猪大肠中合成。色氨酸首先转变为3-吲哚乙酸,随后在大肠杆菌和梭菌的某些种的作用下产生吲哚-3-乙酸,之后乳酸杆菌转变其为粪臭素(3-甲基吲哚)。因此粪臭素最终数量取决于色氨酸的利用度和肠道中菌群组成和活力。色氨酸主要由肠中的黏液细胞转化产生。一部分粪臭素由粪便排出,一部分经过肠道吸收入血。全公猪脂肪中的粪臭素含量很高,不同性别猪粪中粪臭素含量却接近一致。
2.2 粪臭素的代谢
肝脏代谢分为两步。首先加上1 个羟基,也可在二期代谢加上羟基。形成结合物增加了水溶性,促进其经尿液和胆汁排泄,减少脂肪组织的吸收。一期代谢由位于肝细胞内质网上含有血红蛋白的细胞色素P450 同工酶Ⅱ型(CYP2E1,CYP2A)参与。同样,CYP2E1,CYP2A 和醛氧化酶也参与了对粪臭素的氧化代谢,CYP2E 为主要酶。
Diaz(1997)采用肝微粒体技术确定了粪臭素代谢的一系列代谢物,定量化为:3-羟基-3-甲基吲哚(45%),其他为:5-羟基-5-甲基吲哚,6-羟基-3 甲基吲哚,3-甲羟基吲哚,吲哚-3- 甲 醇,和2- 氨 基- 苯 乙 酮。这些代谢产物有些可以作为二期代谢底物加上硫酸盐和葡萄糖醛酸。血浆中高浓度的6-磺基粪臭素可以作为一个指标来判断公猪具有快速代谢和排泄粪臭素的能力,但仍需要进一步研究。苯磺酸转移酶具有热稳定性。对粪臭素代谢中酶类的修饰和表达也许是促进代谢降低膻味的一个有用方法。
粪臭素在肝脏的代谢对于其在脂肪中蓄积有着很大影响。CYP2E1,CYP2A 活力较高的成年公猪脂肪中的粪臭素的含量较低,反之则较高。关于CYP2E1,CYP2A 在粪臭素代谢中作用仍存在争论,也许与研究中采用的动物的年龄不同有关。其他研究发现,粪臭素在青年公猪中的代谢与成年公猪有所不同。
3 雄烯酮、粪臭素和动物的激素状态
雄烯酮的合成与其他睾丸激素一样,受到下丘脑——垂体——性腺轴的控制。因此,在青春期雄烯酮与其他睾丸类固醇类激素数量快速增加。随着睾丸类固醇增加,粪臭素的量也在增加。可能是睾丸类固醇对肝粪臭素代谢加以调控。雌激素含量与粪臭素的水平也存在正相关联系。
体外实验发现,雄烯酮是一个潜在的对CYP2E1 CYP2A 的抑制子。肝脏中CYP2E1 的启动子受到转录因子COUP-TF1 和HNF-1 的活化。Sinclair(2006)发现活化组成型雄甾醇受体(CAR)、孕酮X 受体(PXR)和法尼醇X 受体(FXR)等核受体的配体增加了睾丸间质细胞中SULT2A1的表达。Gillberg(2006)发现原代猪肝脏细胞CAR 受体的活化增加了CYP2A 的活力。因此,似乎粪臭素与雄烯酮的代谢途径受到这些核受体的共同调节。也许是雄烯酮的一些代谢产物与这些核受体结合而影响到粪臭素代谢相关酶的基因表达。

图2 阉割
4 对膻味的调控
4.1 免疫阉割对雄烯酮生物合成的修饰
免疫阉割可以消除促性腺激素(GnRH),减少睾丸类固醇的产生,除了可以减少雄烯酮的合成,还可以减少生殖器官的尺寸、精子数目和进攻性。此外,粪臭素和吲哚的含量也可以下降。辉瑞公司的ImprovacTM疫苗可以进行2 次免疫,2 次之间间隔4 周以减少膻味。免疫阉割抑制了睾丸类固醇的产生,使得肝脏中对粪臭素的代谢活力提高,减少脂肪组织中粪臭素蓄积。同时IGF-1的含量下降,也减少了粪臭素的合成。因此通过免疫阉割,既使肉品质提升又可生产全公猪肉。当进行圈养时,宰前的体重过大而不易免疫操作,因此在第2 次免疫注射和屠宰之前有更长的间隔期是最好的。免疫阉割的一个负面影响导致内源性类固醇下降,猪的生长率和瘦肉率下降。从安全的角度考虑,理想的疫苗注射一次最好。
4.2 通过日粮对粪臭素合成和代谢调控
总的来说,脂肪中的粪臭素的量依赖于肠道中产生的数量、肠道停留时间、小肠吸收和肝脏的代谢。前三者通过调节日粮组成可以加以调控,目前对这个日粮组分的模型正处于研究之中。以前研究发现,日粮中一定的碳水化合物影响着肠道中的微生物区系和肠道的功能,反过来又诱导肠道中产生粪臭素的变化。未消化的碳水化合物可以影响增加粪的湿重和干重,减少粪便在肠道停留时间,减少大肠对粪臭素的吸收。肠道吸收粪臭素量依赖于在水中溶解度和肠道停留时间,然而人们对粪臭素从肠道吸收的机制知之甚少。
菊苣多糖和粗制土豆淀粉(RPS)在猪的日粮中可降低组织和粪中的粪臭素的水平。碳水化合物经肠道内微生物区系加以发酵利用,具有对动物保健和调控肉品的潜在功能。先前研究认为,饲喂RPS 减少吲哚是因为抑制了大肠中细胞凋亡,减少了对色氨酸的利用。RPS对同种动物的粪臭素和吲哚产生影响不同,说明了RPS 除了影响凋亡之外,还影响到其他因素。这些因素也许影响了粪臭素的合成,而对于吲哚的影响甚少。
日粮可以影响到肝脏中CYP 的同工酶,后者随即又影响底物代谢。然而,日粮与粪臭素之间的关系并没有确定。Zamratskaia(2005,2006) 报 道全公猪饲喂RPS 或者不饲喂,CYP2E1和CYP2A 的活力是相同的。Whiteer(2004)报道含有菾菜根的日粮增加了CYP2E1 的表达水平。
4.3 提前宰杀
全公猪在年轻时及体重较低时宰杀可以减少膻味。因不同品种的青春期不同,即使同一品种的不同个体之间,造成的研究结果也不相同。Zamratskaia(2005)研究发现90 kg 和115 kg 屠宰的猪雄烯酮没有差别。Chen(2007)发现在体重较大的猪体内脂肪中的雄烯酮较高。Nicoke(2007)发现较重猪脂肪中两种膻味物质的含量较高。Aldalfaixan(2005)研究杜长大三元组合猪在75 kg 时宰杀,并不能完全得到消除膻味的猪肉。这些研究提示降低屠宰活重并不能完全消除膻味。

图3 猪群
5 未来的研究方向
5.1 膻味化合物
雄烯酮和粪臭素在组成膻味中特别重要,但膻味的组成不单限于这2 种化合物,其他如:吲哚、雄烷醇、苯甲亚基丙酮和短链脂肪酸等尚需要确定。
还要建立对膻味进行有效和一致感觉评估的标准。目前尚缺乏膻味的阈值标准,尤其是雄烯酮。通常使用粪臭素的阈值为0.2~0.25μg/g,雄烯酮为0.5~1.0μg/g。雄烯酮阈值这么大的变化是因为不同消费者对于膻味的感觉能力不同。因此雄烯酮的阈值范围应该不依赖于人群的差异和检测方法的不同。为了确定膻味感觉阈值,最根本是要有一个基于标准检测程序的检测结果。
5.2 膻味的生物化学
粪臭素在大肠中的合成已经进行了很多研究,但粪臭素产生的微生物机制仍然知之不多。研究产生高或低粪臭素猪的肠道微生物区系,有助于了解色氨酸在大肠内是怎样降解,以便于我们进一步操控微生物环境以减少粪臭素的产生。CYP2E1 和CYP2A 2 种酶在粪臭素的代谢中起着关键作用,要进一步调查各种睾丸类激素对粪臭素在肝脏代谢影响。此外,雄烯酮形成磺基结合物及其对于雄烯酮在脂肪中消除和积累也应进一步研究。
5.3 膻味的遗传学
要慎重采用在其他动物中评估动物性成熟的办法来评估公猪膻味的遗传潜能。可以从现有商用品种中采集大量数据来解释遗传标记SNPs(single nucleotide olymorphisms,单核苷酸多态性)与膻味之间的联系。更重要的是确定膻味数量性状基因座,确定这些基因座可以用来比对染色体上膻味基因的位置。采用具有相同类固醇产生潜力,而表现型具有高或低膻味的猪来进一步对基因表达做研究,以确定候选基因。最后,也应对膻味物质代谢整合过程研究进行支持,这些研究可以发现更多的候选基因。
5.4 免疫阉割
免疫阉割与全公猪的成本效益分析尚未有人进行对比。进行此实验要消耗人工、材料和大量的免疫费用。免疫阉割的时间是否可以比现在推荐的时间更早一些?免疫阉割对猪肉品质影响的研究甚少,仅有几篇文献报道了保水能力和色泽。
5.5 膻味检测方法
屠宰场内快速检测膻味物质面临着挑战。现有技术要么处于开发阶段,要么需要高素质技术人员来操作。理想的技术应该在较短时间内(几秒~几分钟)就应得到结果。很多情况下,采样的时间耗去了实验的大部分时间。另外,现有的检测方法昂贵,能效比决定着业界是否采用新的检测技术。
理想技术的检测结果是:区分可接受样品和不可接受样品应该是百分之百的正确。鉴于不同国家的市场情况,低的假阳性可以接受,但假阴性必须为零。指纹印迹检测技术仍然有太高的假阴性(5%~10%)。目前没有一种适用于各种屠宰条件的技术。还需要开发相应的工作软件,要考虑到投入产出比、自动化、高通量,取样简单易行。
本文摘编自:
K. Lundstrom,K.R.M atthews,J.E.Haugen.Pig meat quality from entire males.Animal,2009,11(3):1497-1507.
G.Zamaratskaia,, E. J. Squires.Biochemical,nutritional and genetic effects on boar taint in entire male pigs.Animal,2009,11(3):1508-1521.
