中国中北部青檀的AFLP分析
2016-07-15朱翠翠张林孙忠奎王长宪山东农业大学林学院山东泰安7000泰安市泰山林业科学研究院山东泰安7000泰安时代园林科技开发有限公司山东泰安7000
朱翠翠,张林,孙忠奎,王长宪(山东农业大学林学院,山东泰安7000;泰安市泰山林业科学研究院,山东泰安7000;泰安时代园林科技开发有限公司,山东泰安7000)
中国中北部青檀的AFLP分析
朱翠翠1,张林2,孙忠奎3,王长宪2
(1山东农业大学林学院,山东泰安271000;2泰安市泰山林业科学研究院,山东泰安271000;3泰安时代园林科技开发有限公司,山东泰安271000)
摘要:为揭示中国中北部青檀的遗传多样性,笔者从贵州、北京、甘肃、江苏、安徽、山东、河南7省12个种群收集了31份青檀样品,对其进行AFLP分析。利用从26对AFLP引物组合中筛选出的6对引物对样本DNA进行扩增,共产生223条谱带,其中211条为多态性条带,比率达94.26%。结果显示中国中北部的青檀材料之间存在较高的遗传多样性,且AFLP技术可以有效地对其进行分析。31个样品的遗传相似系数在0.6413~0.9193之间,平均为0.7489,聚类分析结果显示样品首先按照种群聚在一起,但也有交叉。
关键词:青檀;聚类分析;AFLP;遗传多样性
0 引言
青檀(Pteroceltis tatarinowii Maxim.),又名翼朴,为榆科(Ulmaceae)青檀属(Pteroceltis)落叶乔木,中国的特有植物,在中国植物红皮书中被列入国家三级重要保护对象,是中国特有的纤维树种和钙质土壤的重要指示植物。在中国皖、陇、贵、苏、鲁、鄂、湘等19省均有生长。青檀适应性强,耐旱耐湿、耐盐碱,是石灰岩裸露岩石荒山造林的重要树种。青檀作为中国重要的乡土树种和绿化树种,在园林景观建设过程中,与乔、灌、草相结合,不仅可以美化环境,还可以改善城市小气候、维持城市生态系统的平衡,其叶可作为高级营养型饲料添加剂,其檀皮制作的宣纸一直被书法家和画家视作珍品,其木材坚硬致密也可以用来制作家具,具有很高的应用价值[1-4]。
由Vos等[5]发明的AFLP(Amplified fragment length polymorphism,扩增片段长度多态性)分子标记技术是通过不同长度的限制性内切酶片段检测DNA多态性的一种分子标记方法。与其他分子标记方法相比,AFLP技术快速、稳定、高效,并且DNA用量少,检测效率高[6-8]。该技术自出现起就被广泛应用于遗传多样性的研究,如银杏[9]、枣树[10]、秋子梨[11]等。研究遗传多样性对于植物资源利用、保护及品种选育等有重要意义[12]。
由于人为的破坏和大范围的人工繁殖栽培,野生青檀日益减少,导致青檀遗传多样性日益减少。但目前,有关青檀遗传多样性的研究有零星报道,李晓红等[13]用ISSR对来自广东、福建、浙江、安徽等27个自然种群的青檀样品进行了初步分析,林巧红[14]对山东灵岩寺、佛峪以及枣庄青檀寺野生青檀资源进行了调查与评价等。笔者对来源于北京、山东、安徽等地12个种群的青檀资源进行AFLP分析,意在揭示中国中北部青檀的亲缘关系和遗传多样性,为青檀资源保护、品种改良奠定基础。
1 材料与方法
1.1植物材料
供试样品来自于北京昌平、甘肃文县、安徽泾县、江苏南京、贵州贵阳、山东济南灵岩寺等地12个种群。在各个种群内随机选取正常、无病害的植株嫩叶进行采集,放入装有变色硅胶的塑料袋中进行快速干燥。表1为样品采集地点和编号。

表1 样本材料编号及来源
1.2试验方法
1.2.1DNA提取改良CTAB法提取基因组DNA并检测纯度和浓度。
1.2.2AFLP试验
(1)酶切。对所选样品采用EcoRⅠ/MseⅠ双酶切(ferments酶):
酶切体系(20 μL)为:10×Buffer tango 4 μL (fermentas);EcoRⅠ(10 U/μL)0.4 μL;MseⅠ(10 U/μL)0.4 μL;模板DNA约200 ng;补ddH2O至20 μL。
酶切反应条件为37℃,3 h,65℃,3 h。
(2)连接。连接体系(20 μL)为:10×T4 Buffer 2 μL;EcoR I Adaptor(50 μmol/L)1 μL;MseⅠAdaptor (50 μmol/L)1 μL;T4 ligase 0.4 μL(5 U/μL)酶切产物10 μL;补ddH2O至20 μL。16℃过夜。
(3)预扩增。反应体系:10×Buffer 2 μL;10 mmol/L dNTPs0.4μL;5 U/μLTaq酶0.2μL;E000.5μL(20μmol/L);M000.5μL(20μmol/L);连接模板2μL;加ddH2O至20μL。
PCR扩增程序为:94℃预变性3 min;94℃变性45 s;50℃复性45 s,72℃延伸1 min,26个循环。
(4)选择性扩增。反应体系:10×Buffer 2 μL;10 mmol/L dNTPs 0.4 μL;5 U/μL Taq酶0.2 μL;Eprimer1 μL(20 μmol/L);Mprimer1 μL(20 μmol/L);模板DNA2 μL;加ddH2O至20 μL。
PCR扩增程序为:95℃预变性5 min;95℃变性35 s,65℃复性35 s(每循环降低0.7℃),72℃延伸1 min,12个循环;94℃变性30 s,56℃复性30 s,72℃延伸1 min,23个循环。
(5)变性电泳。PCR产物加上上样缓冲液,94℃变性10 min后进行PAGE分析,银染后观察。
1.2.3数据分析利用Quantity One软件将31个样品、6对引物的AFLP PAGE电泳图根据无带和有带情况转化为0、1数据矩阵。采用POP软件计算个体遗传距离和相似系数,并使用NTSYS软件进行UPGMA聚类分析。
2 结果与分析
2.1样品AFLP多态性及扩增效率
通过筛选,笔者在26对AFLP引物组合中共筛选出6对多态性引物,信号强度一致、条带分布均匀。6对引物共产生223条带(见表2),其中211条多态性条带,所占比例(PPB)达94.62%,平均每对产生35.2条。引物组合E35M83、E40M55、E41M64、E43M54、E76M47的多态性都在90%以上,引物组合E35M83最高达98.36%,引物组合E86M85最低为75%。图1显示了12个青檀种群31个样品的遗传图谱,几乎所有种群的样品都能被区分,这说明青檀具有丰富的遗传变异。AFLP技术可以有效地用于中国中北部青檀的遗传多样性和亲缘关系分析,且多态性和可重复性都比较高。

表2 引物名称及多态性条带比较

图1 引物E40M55的AFLP图谱
2.2相似性分析
图2显示了31个青檀样本的遗传相似系数。结果显示,31个青檀样本之间的遗传相似系数在0.6413~0.9193之间,表明中国中北部青檀样品之间存在一定的遗传差异。北京檀峪村样品18、19之间最大为0.9193,说明两者变异最小,遗传距离最近。而檀峪村样品18和文县样品26之间最小为0.6413,说明两者变异最大,遗传距离最远。对于同一分布区的青檀样品而言,来自北京的样品18和19之间遗传相似系数最大0.9193,来自甘肃文县的样品26和30遗传相似系数最小0.6906,这说明同一个种群的青檀样品之间也存在一定程度上的遗传差异性。
2.3不同种群青檀的聚类分析
如图3聚类图显示,以0.740为域值,供试青檀样品可分为3类。第Ⅰ类包括青阳(1号)到文县(30号)共17个样品,安徽(青阳、泾县、淮北大方寺)、南京(燕子矶)、山东(曲阜、济南檀抱泉)以及甘肃文县的样品被归到第Ⅰ类。第Ⅱ类包括北京、济南灵岩、河南的样品共11个。山东泰安东平(31号)被归到第Ⅲ类。以0.755为域值,供试青檀样品分为5亚类。第Ⅰ亚类包括安徽(青阳、淮北大方寺、泾县)的7个样品和南京燕子矶的3个样品以及山东曲阜的1个样品。甘肃文县的5个样品以及檀抱泉的1个样品构成第Ⅱ亚类。来自贵州花溪的5个样品、北京的3个样品、灵岩的4个样品构成第Ⅲ亚类。河南南召(17号)和泰安东平(31号)各成一类。东平(31号)、河南(17号)样品过少,聚类结果有待进一步试验验证。从聚类结果上看,同一种群内青檀具有一定的遗传多样性,而不同种群青檀的遗传关系远近在地域范围上呈现一定的相关性,但并不是很明显,这说明青檀种质资源遗传距离的远近与地理距离并没有显著相关关系。
3 讨论
笔者利用AFLP分子标记在一定程度上有效地揭示了中国中北部青檀的遗传多样性。把分子水平以及生态环境结合起来评价物种的遗传多样性对于人们合理地利用和保护中国青檀资源具有重要意义[15-17]。到现在为止,有关青檀遗传多样性的工作开展不多,限制了青檀种质资源的遗传改良和保护工作。Chai等[18]采用ISSR技术对皖、豫、苏、鲁4省5个种群的样品进行分析,显示青檀种群内和种群外都具有较高的遗传多样性;李建华等[19]对湖北省大贵寺国家森林公园不同海拔高度进行ISSR分析结果表明,遗传关系与海拔相关;林巧红[14]对来自青檀寺、佛峪、和灵岩寺3个种群进行SRAP分析。本研究从贵州、山东、江苏、安徽、北京、河南收集了31份青檀资源,对中国中北部的青檀资源进行遗传多样性分析。AFLP结果显示,6对引物共扩增出223条多态性带,百分比(PPB)达94.62%,说明用AFLP标记可以有效的分析青檀遗传多样性。从结果上看贵阳与北京檀峪村、灵岩寺之间遗传相似性相对较大,这可能和人为活动有关。对中国中北部不同种群青檀样品进行AFLP分析的聚类结果表明,尽管亲缘关系的地域趋势在一定的地域范围内呈现出来,但总的来看,遗传距离的远近并没有显著的地域相关性。Fischer等[20]认为,当遗传漂移存在时,遗传距离的远近和地理距离就没有显著的相关性,只有起主导作用的是基因流时,两者才会呈现出明显的相关性。按Fischer的观点,可以这样推测,长期以来由于青檀具有一定的经济价值,青檀的野生资源遭到人为的过度挖掘而逐渐减少,野生资源日益匮乏,种群也逐渐减少,而产生一定的遗传漂移。青檀的相互引种、生活史类型、繁殖方式、交配系统以及基因流,都可能是形成青檀种群内部和青檀种群间遗传变异差别的主要原因。
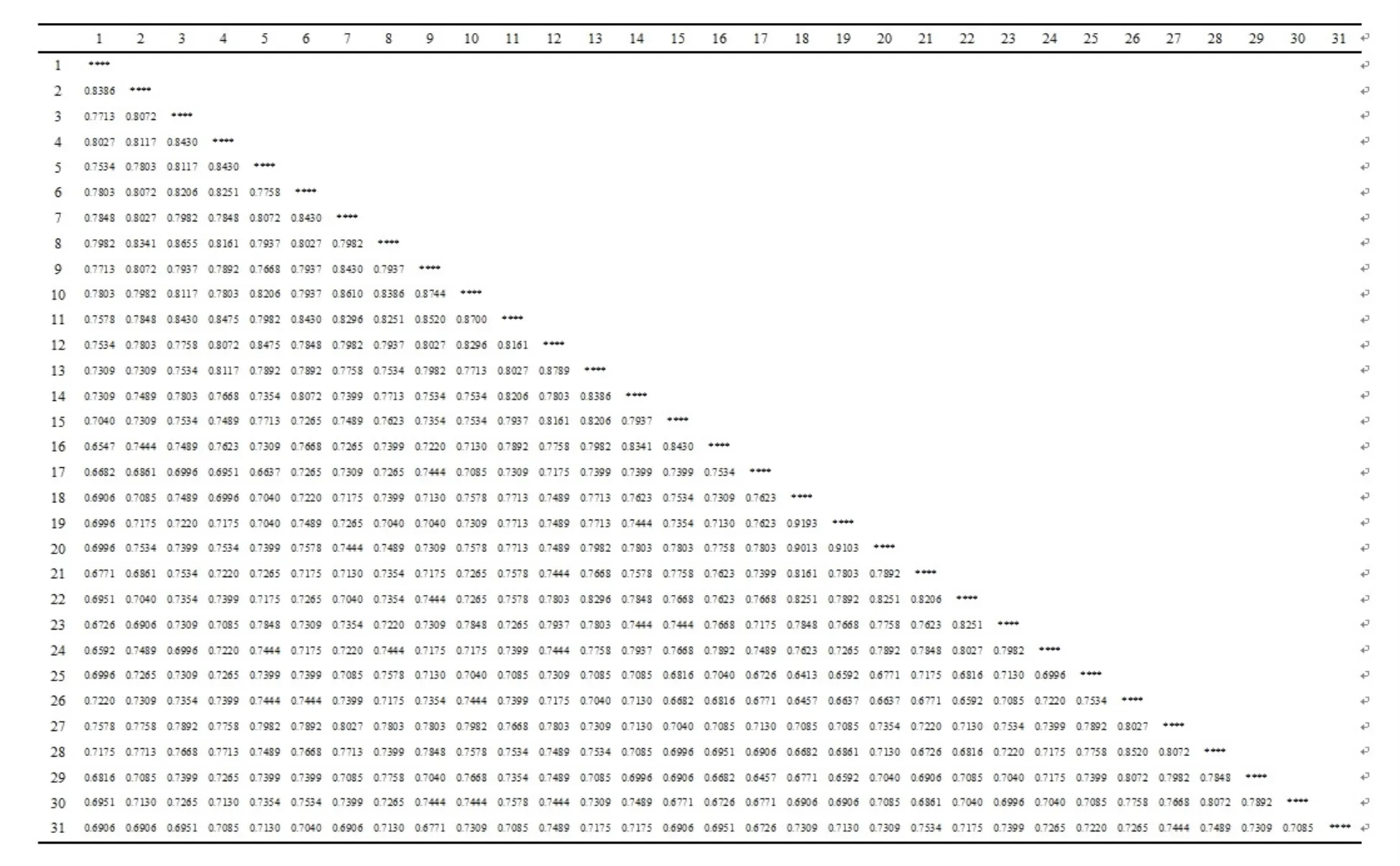
图2 不同种群青檀31个样品遗传相似系数

图3 不同地区青檀聚类图
参考文献
[1]张天麟.园林树木1600种[M].北京:中国建筑工业出版社,1988: 115.
[2]陈有民.园林树木学[M].北京:中国林业出版社,2002:328.
[3]张林,王峰,孙忠奎,等.青檀多倍体诱导试验初报[J].中国农学通报, 2015,31(13):1-4.
[4]许冬芳,崔同林.青檀的开发利用[J].中国林副特产,2005(3):64.
[5]Vos P R, Hogers M. AFLP: a new technique for DNA fingerprinting [J].Neu Res.,1995,23:4407-4414.
[6]Zabeau M, Vos P. Selective restriction frsgment amplification, a meathod for DNA fingerprinting. European Patent Application 94202629.7(publication No.05348Al)[P].Paris: European Patent Office,1993.
[7]Vos P, Hogers R, Bleeker M, et al. AFLP: a new technique for Dna fingerprinting[J].Nucleic Acids Res,1995,23(21):4407-4414.
[8]王斌,翁曼丽.AFLP的原理及其应用[J].杂交水稻,1996(5):27-30.
[9]王利.银杏种质资源遗传多样性的AFLP研究[D].泰安:山东农业大学,2006.
[10]乔勇,赵锦,杨海旭,等.21个枣品种系的AFLP指纹分析[J].植物遗传资源学报,2009,10(2):205-210.
[11]安萌萌,刘畅,何天明,等.黑龙江省野生秋子梨群体遗传结构的荧光AFLP分析[J].植物遗传资源学报,2014,15(4):728-733.
[12]关萍,石建明,陈放.不同分布区天麻的AFLP分析[J].植物遗传资源学报,2013,14(1):70-77.
[13]李晓红,张慧,王德元,等.我国特有植物青檀遗传结构的ISSR分析[J].生态学报,2013,33(16):4892-4901.
[14]林巧红.山东省珍稀物种青檀的种植资源调查与评价[D].泰安:山东农业大学,2013.
[15]胡尊红,郭鸿彦,胡学礼,等.大麻品种遗传多样性的AFLP分析[J].植物遗传资源学报,2012,13(4):555-561.
[16]李凤鸣,张华新,杨秀艳.省沽油群体遗传多样性的AFLP分析[J].植物遗传资源学报,2012,13(1):130-133.
[17]李明,高宝嘉,张静洁.承德光秃山不同海拔油松居群遗传多样性与生境因子关联研究[J].植物遗传资源学报,2012,13(3):350-356.
[18]Chai X Y, Chen S L. Xu W. Using inter-simple sequence repeat markers to analyze the genetic structure of natural Pteroceltis tatarinowii populations and implications for species conservation[J]. Plant Syst Evol.,2010,285:65-73.
[19]李建华,李雪松,田春元,等.大贵寺国家森林公园野生青檀居群的遗传多样性[J].生态环境学报,2011,20(12):1799-1804.
[20]Fischer M, Husi R, Prati D, et al. RAPD variation among and within small and large population of the rare clonal plant Ranunculus reptans(Ranunculaceae)[J].Am J Bot.,2000,87(8):1128-1137.
AFLP Analysis of Pteroceltis tatarinowii in the North of Central China
Zhu Cuicui1, Zhang Lin2, Sun Zhongkui3, Wang Changxian2
(1College of Forestry, Shandong Agricultural University, Tai’an 271010, Shandong, China;2Taishan Forestry Science Institute, Tai’an 271000, Shandong, China;
3Tai’an Shidai Technology Development Limited Company, Tai’an 271000, Shandong, China)
Abstract:In order to study the genetic diversity of Pteroceltis tatarinowii in the north of central China, 31 samples were collected from 12 populations from 7 provinces(Guizhou, Beijing, Gansu, Jiangsu, Anhui, Shandong and Henan)and AFLP analysis was conducted. A total of 223 bands were generated from 6 primer combinations screened from 26 primer combinations and 211 of those bands showed polymorphism, the polymorphic ratio(PP)was up to 94.26%. The results showed that a high genetic diversity was existed among Pteroceltis tatarinowii samples from the north of central China and AFLP analysis was effective. The genetic similarity coefficient was in the range of 0.6413 to 0.9193, with an average value of 0.7489. And the results of cluster analysis showed that the samples were clustered together according to the population, but there were intersections between the populations.
Key words:Pteroceltis tatarinowii; Cluster Analysis; AFLP; Genetic Diversity
中图分类号:S688
文献标志码:A论文编号:cjas16010002
基金项目:2014年山东省农业良种工程项目“青檀种质创新利用与新品种培育”(鲁科字[2014]96号)。
第一作者简介:朱翠翠,女,1991年出生,德州齐河人,硕士研究生,研究方向:园林植物。
通信地址:271000山东省泰安市泰山区岱宗大街61号山东农业大学北校区林学院404,E-mail:zhucuicuitt@163.com。 271000山东省泰安市泰山林业科学研究院,E-mail:changxianwang@163.com。
通讯作者:王长宪,男,1959年出生,山东平阴人,研究生导师,研究员,研究方向:园林植物遗传育种。
收稿日期:2016-01-04,修回日期:2016-03-05。
