原生动物对白蚁食性的影响研究
2010-09-04徐忠东张有森陶瑞松
徐忠东, 张有森, 陶瑞松, 张 雁
(1.合肥师范学院生命科学系,安徽合肥230061;2.合肥白蚁防治研究所,安徽合肥230000)
原生动物对白蚁食性的影响研究
徐忠东1, 张有森2, 陶瑞松1, 张 雁1
(1.合肥师范学院生命科学系,安徽合肥230061;2.合肥白蚁防治研究所,安徽合肥230000)
本文介绍了低等白蚁的原生动物分布类型,阐述了原生动物对白蚁食性的影响,探讨了原生动物帮助白蚁消化代谢的机理和在共生体系的作用。
原生动物;白蚁;分类;食性
白蚁属昆虫纲等翅目的社会性昆虫,不完全变态,世界性发布,主要发布在热带、亚热带。白蚁对人类生活和经济影响重大,其危害房屋建筑、江湖堤坝、农林植物、电缆、交通设施等相当严重,但对人类也有益,如营养价值较高,是森林的“清道夫”,可加速物质循环,改变土壤理化性质[1]。
1 白蚁分类
白蚁在自然界广泛分布,大约2600多种[2][3],我国有522种[1]。不同学者对白蚁分类观点存在差异,主要从形态结构、表皮碳氢化合物、食性、肠道共生微生物、DNA序列分析等方面划分。Holmgren将白蚁分为4个科,Snyder把它分成7个科[2]。按Kumar Krishna的分类,分为6个科,即澳白蚁科(Mastotermitidae Silvestri,1909)、木白蚁科(Kalotermitidae Enderlein,1909)、草白蚁科(Hodotermitidae Snyder,1925)、鼻白蚁科(Rhinotermitidae Light,1921)、齿白蚁科(Serritermitidae Emerson, 1965)、白蚁科(Termitidae Westwood 1840),其中澳白蚁科、木白蚁科、草白蚁科、鼻白蚁科为低等白蚁,我国有木白蚁科、草白蚁科、鼻白蚁科、白蚁科共43属522种[1]。
生态环境对白蚁的多样性产生影响,根据其分布和对纤维素、木质素分解能力的差异,分为食木类、食菌类、食土类。根据筑巢地点,也可再进行区分。
2 白蚁中原生动物分布类型
白蚁之所以能有效利用食物中的木质纤维素物质,与其肠道共生微生物有很大关系。白蚁肠道一般分为前肠、中肠和后肠,后肠相当发达,约占肠道总容积的80%,由肠道瓣前节、肠道瓣、囊形胃、结肠和直肠组成,为一条粗细不均的管道,囊形胃特别膨大。后肠是厌氧的消化器,有大量的共生微生物,可降解纤维素与半纤维素,形成白蚁可吸收的短链脂肪酸[1][4]。
高等白蚁白蚁科共生微生物中很少有原生动物,其消化纤维素和半纤维素依靠唾液腺或中肠上皮细胞分泌的内源性纤维素酶,有的是从巢体上共生真菌处获得外源性纤维素酶。
低等白蚁的肠道特别是后肠含有多种多样的微生物,其中原生动物可占白蚁总体重的1/7—1/3,原生动物主要是鞭毛虫,已发现有434种[5],这些鞭毛虫属于鞭毛纲(mastigophorea)中的3个目,即毛滴虫目(Trichomonadida)、超鞭虫目(Hypermastigida)和锐滴虫目(Oxymonadida)[6][7]。它们都没有线粒体,毛滴虫目和超鞭虫目的鞭毛虫是较原始的原生动物。这些鞭毛虫有一定的宿主专一性,分布于不同的低等白蚁(表1)[5]。
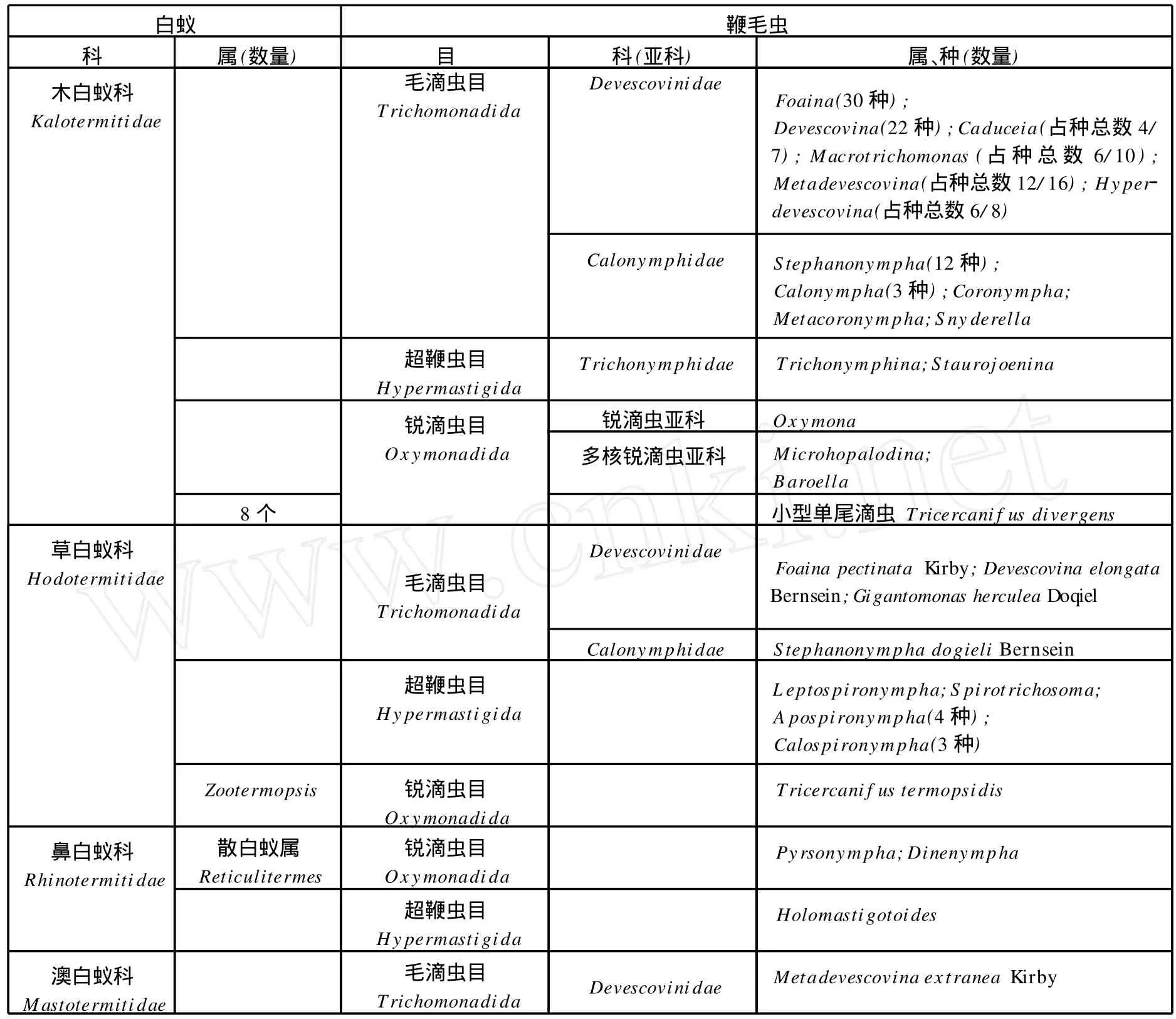
表1 低等白蚁中原生动物分布
3 原生动物对白蚁食性的影响
在低等白蚁中,共生的原生动物非常重要,如果没有它们,白蚁就不能消化木质纤维素。人们通过通氧(3.5大气压)或较高的温度(如36℃中24h)处理白蚁,杀死肠道内鞭毛虫,再继续饲喂木材,白蚁2周后死亡[1][8]。当使消除了鞭毛虫的白蚁重新感染上鞭毛虫后,其存活时间明显较未重新感染鞭毛虫的白蚁长[9]。这说明共生的原生动物等微生物决定了其宿主白蚁的食物类型。白蚁与其肠道共生微生物在长期进化中形成了其摄食需求,从白蚁肠道内原生动物分布也可看出不同的白蚁食性上有所差异。
当改变白蚁食物类型,其肠道原生动物也产生变化。Peterson等发现亚洲地下白蚁 Reticulitermes f lavipes(Kollar)(Isoptera:Rhinotermitidae)食用火烧后的木头,造成体内原生动物明显减少[10]。白蚁 Coptotermes f ormosanus.后肠共生三种原生动物 Pseudotrichonympha grassi,Holomastigotoides hartmanni,和 S pirotrichonympha leidyi,Tanaka等用日本红松木屑、纤维素、纤维二糖、葡萄糖人工喂养低等白蚁 Coptotermes f ormosanus.30天后,P.grassi和 H.Hartmanni消失,仅有少量 S.leidyi存在,说明 P.grassi和 H. Hartmanni在白蚁消化利用高分子量碳水化合物起重要作用[11]。
Nakashima等通过聚丙烯酰胺凝胶电泳法酶谱分析,C.f ormosamus拥有两个独立的纤维素消化系统,一个白蚁中肠自身的纤维素酶,另一个是白蚁后肠共生的原生动物产生的[12]。
Maistrello 等 用 disodiumoctaborate tetrahydrate、努特卡酮或两者混合物处理木头,喂养白蚁 Coptotermes f ormosanusShiraki,结果表明,在白蚁消化纤维素起重要作用的鞭毛虫 Pseudotrichonympha grassiiKoidzumi几乎全部消失[13]。
Kard试验发现,在生物鉴定法中,硼酸与土壤混合物引起地下白蚁明显死亡。在选择性实验中,将亚洲白蚁和台湾白蚁暴露于浓度为0.05,0.25, 0.50,1.00,2.00,and 4.00 percent Al(wt:wt)硼酸与土壤混合物12周,两种白蚁肠中原生动物没有明显下降,无剂量依赖性[14]。
白蚁消化食物中纤维素是利用自身和体内共生微生物分泌的纤维素酶共同作用。将纤维素水解为葡萄糖所需要的酶包括内切β-1,4-葡聚糖酶、外切葡聚糖酶(纤维二糖酶,外切β-1,4-葡聚糖酶)和β-葡萄糖苷酶等。在低等白蚁中,纤维素酶主要来源于原生动物。如在台湾乳白蚁(Coptotermes f ormosanus)中,外-纤维二糖水解酶87%的活性来自后肠的鞭毛虫[5]。应用PCR和原位杂交等技术从栖北散白蚁、澳白蚁和台湾乳白蚁肠道共生鞭毛虫中克隆出了,如葡糖水解酶45家族[15][16]、水解酶 GHF7[17]和β-1,4-葡聚糖酶[18]等不同的纤维素酶基因。
日本学者本乡裕一等对白蚁肠道鞭毛虫 Trichonympha agilis研究发现,其细胞内存在共生细菌Rs-D17,属 Termite Group 1(TG1)门,基因组极度缩小,拥有15种氨基酸、嘌呤和嘧啶的生物合成系统,合成白蚁和原生动物无法合成的一些含氮化合物,供给原生动物和白蚁。这对于研究原生动物在白蚁肠道共生作用及形成白蚁、原生动物、细菌多重共生系的机理具有重要意义[19]。
4 讨论
虽然目前对低等白蚁肠道中存在的鞭毛虫及与白蚁分类的关系有一些研究,但对其他原生动物如纤毛虫了解很少。对白蚁原生动物研究仍然主要是形态方面研究,其培养存在困难,仅有少量白蚁原生动物培养成功,不利于其准确分类和对白蚁分类的支持。随着分子生物学技术发展,如DNA指纹图谱技术,将有利于深入了解不同类型、不同食性、不同品系白蚁的原生动物类型;探讨细菌、原生动物、白蚁共生机制,筛选具有较高活性纤维素酶的原生动物和开发高活性纤维素酶基因工程菌株创造条件,也为研制纤维素酶抑制剂防治白蚁提供手段。
[1] 黄远达.中国白蚁学概论[M].湖北:科学技术出版社,1999.
[2] 戴华国,李小鹰,张红兵.白蚁分类方法述评[J].昆虫知识, 2004,41(2);20-23.
[3] 曹迪,赵凯,平文祥,等.高等白蚁 Macrotermes subhyalinus消化道的研究进展[J].高师理科学刊,2008,28(3):63-65.
[4] 秦伟,王维平.白蚁消化道及肠道微生物研究简述[J].江西植保,2007,3 0(2):68-69.
[5] 杨红,彭建新,刘凯于.低等白蚁肠道共生微生物的多样性及其功能[J].微生物学报,2006,46(3):496-499.
[6] Radek R.Flagellates,bacteria,and fungi associated with termites:diversity and function in nutrition-a review.Ecotropica, 1999,5:183-196.
[7] 李珺,张贞华,李桃生.低等白蚁肠道内原生动物的分布及其进化学意义[J].白蚁科技,1999,16(2):1-7.
[8] Radek R.Flagellates,bacteria,and fungi associated with termites:diversity and function in nutrition—a review[J].Ecotropica,1999,5:183-196.
[9] Yamin MA.Cellulose metabolism by flagellaTrichonymphs sphaerticafrom a termite is independent of endosymbiotis bacteria[J].Science,1981,211:58-59.
[10] Peterson,C J Gerard,P D Wagner,T L Charring does not affect wood infestation by subterranean termites[J].Entomologia Experimentalis et Applicata,2008,126(1):78-84.
[11] Tanaka,H Aoyagi,H Shina,S et al Influence of the diet components on the symbiotic microorganisms community in hindgut ofCoptotermes f ormosanusShiraki[J].Applied Microbiology and Biotechnology 2006,71(6):907-917.
[12] K.Nakashima H.Watanabe H.Saitoh Dual cellulose-digesting system of the wood-feeding termite,Coptotermes f ormosanusShiraki[J]Insect Biochemistry and Molecular Biology, 2002,32(7):777-784.
[13] Maistrello L Henderson G Laine RA.Effects of nootkatone and a borate compound on formosan subterraneantermite (Isoptera:Rhinotermitidae)and its symbiont protozoa[J]. Journal of entomological science,2001,36(3):229-236.
[14] Kard BM.Detrimental effects of boric-acid-treated soil against foraging subterranean termites(Isoptera:Rhinotermitidae)[J].Sociobiology,2001,37(2):363-378.
[15] Ohtoko K,Ohkuma M,Moriya S,et al.Diverse genes of cellulase homologues of glycosyl hydrolase family 45 from the symbiotic protist in the hindgut of the termiteReticulitermes speratus[J].Extremophiles,2000,4:343-349.
[16] Li L,Froehlich J,Pfeiffer P,et al.Termite gut symbiotic Archaezoa are becoming living metabolic fossils[J].Eukaryotic Cell,2003,2:1091-1098.
[17] Nakashima K,Watanabe H,Saitoh H,et al.Dual cellulosedigesting system of the wood-feeding termites,Coptotermes f ormosanusShiraki[J].Insect Biochem Mol Biol,2002,32: 777-784.
[18] Inoue T,Moriya S,Ohkuma M,et al.Molecular cloning and characterization of a cellulase gene from a symbiotic protist of the lower termite,Coptotermes f ormosanus[J].Gene,2005, 349:67-75.
[19] 汐田真理.日本研究人员解读白蚁肠内共生菌的全基因组取得新突破[J]生物产业技术,2008,06/www.biobusiness. com.cn.
Study on Effect of Protozoa on Termite Ingestion
XU Zhong-dong1, ZHANG You-sen2, TAO Rui-song1, ZHAN G Yan1
(1.Department of L if e Science,Hef ei N ormal University,Hef ei230061,China; 2.Hef ei Institute ofTermite Prevention,Hef ei230000,China)
This paper introduces the distribution pattern of protozoa in lower termite gut,and discusses the effect of protozoa on termite ingestion,and the digestion mechanism of protozoa for termite and contribution of it in symbiotic association.
protozoa;termite;classification;ingestion
Q968
B
1674-2273(2010)06-0100-04
2010-07-20
安徽省高校自然基金项目(2006KJ194B)
徐忠东(1965-)男,安徽桐城人,合肥师范学院生命科学系教授。研究方向:原生动物细胞分子生物学。
