牡丹非特异性脂质转移蛋白基因PsLTP的生物信息学分析
2024-02-02问荣荣苏钰高泽芳徐梦宇何雯雯王步勇
问荣荣 苏钰 高泽芳 徐梦宇 何雯雯 王步勇



摘要 [目的]了解牡丹非特异性脂質转移蛋白基因LTP的基因特征及其编码蛋白的生物学特性,探索其生物功能。[方法]运用Blast、ORF-Finder、ProtParam、SignalP 4.1、ProtScale和TMHMM 2.0 Server等生物信息学软件,分析牡丹nsLTP基因序列及其编码蛋白的基本理化性质、信号肽、磷酸化位点、保守结构域等,并运用DANMAN、MEGA软件进行多序列比对和系统发育树构建。[结果]牡丹nsLTP基因(NCBI登录号为:JZ840269.1)序列全长为685 bp,开放阅读框长为354 bp,编码蛋白包含117个氨基酸,理论等电点为8.93,为不稳定性蛋白。牡丹nsLTP蛋白为疏水性蛋白,含有信号肽但不含跨膜结构域,NetPho Server预测其含有8个丝氨酸和3个酪氨酸磷酸化位点及5个苏氨酸磷酸化位点。PsnsLTP蛋白具有植物LTP蛋白典型结构,即4个α-螺旋,4对二硫键,1个可结合和容纳脂质分子的疏水结构。多序列比对及系统发育树分析显示,PsLTP蛋白与绒毛烟草LTP蛋白(XP_009608993.1)的相识度62.7%,且进化树聚为同一小分支,遗传距离较近。[结论]通过对牡丹nsLTP基因特性及蛋白结构研究,系统性研究了牡丹nsLTP基因的生物信息学特性,为牡丹抗病抗逆功能基因的研究提供了依据,也为其他植物nsLTP基因研究提供了参考。
关键词 牡丹;非特异性脂质转移蛋白基因LTP;蛋白结构;生物信息学
中图分类号 Q781;S685.11 文献标识码 A
文章编号 0517-6611(2024)02-0087-06
doi:10.3969/j.issn.0517-6611.2024.02.018
开放科学(资源服务)标识码(OSID):
Bioinformatics Analysis of Peony Non-specific Lipid Transfer Protein Gene PsLTP
WEN Rong-rong,SU Yu,GAO Ze-fang et al
(College of Agricultural and Biological Engineering,Heze University,Heze,Shandong 274015)
Abstract [Objective]In order to understand the genetic characteristics of peony non-specific lipid transfer protein gene LTP and the biological characteristics of its encoding protein,and further explore its biological functions.[Method]Bioinformatics software such as Blast,ORF-Finder,ProtParam,SignalP 4.1,ProtScale,and TMHMM 2.0 Server,were used to analyze the basic physicochemical properties,signal peptides,phosphorylation sites and conserved domains of the nsLTP gene sequence. Moreover,DANMAN and MEGA software were used for the multiple sequence alignment and phylogenetic tree construction of nLTP.[Result]The results showed that the total length of peony nsLTP gene (JZ840269.1) was 685 bp,the open reading frame length was 354 bp,the encoded protein contained 117 amino acids,and the theoretical isoelectric point was 8.93,which was an unstable protein. nsLTP protein is a hydrophobic protein with signal peptide but no transmembrane domain. NetPho Server predicted that nsLTP protein contains 8 serine and 3 tyrosine phosphorylation sites and 5 threonine phosphorylation sites. The nsLTP protein has the typical structure of plant LTP proteins including four α-helix,four pairs of disulfide bonds,and a hydrophobic structure that can bind and hold lipid molecules. Multiple sequence alignment and phylogenetic tree analysis showed that PsLTP protein and LTP protein (XP_009608993-1) were 62.7% familiar,and the phylogenetic tree was clustered into the same small branch.[Conclusion]By studying the nsLTP gene characteristics and protein structure of peony,the bioinformatics characteristics of nsLTP gene of peony were systematically studied,which provided a basis for the study of disease resistance and stress resistance gene of peony,and also provided a reference for the study of nsLTP gene of other plants.
Key words Peony;Non-specific lipid transfer protein gene LTP;Protein structure;Bioinformatics
作者简介 问荣荣(1987—),女,山西大同人,讲师,博士,从事植物病理分子研究。*通信作者,副教授,博士,从事林木病理学及分子生物学研究。
收稿日期 2023-02-12
脂质转移蛋白(lipid transfer protein,LTP)是一类广泛存在于动物、植物和微生物中的碱性小分子脂质结合蛋白,因被认为在体外膜间进行脂质转移而命名。动物体内的脂质转移蛋白有特异性与非特异性之分,而植物体内的脂质转移蛋白,因其脂转移活性具有广泛性和不确定性,被称为非特异性脂质转移蛋白(non-specific lipid transfer protein,nsLTP)。植物nsLTP是一类在高等植物中普遍存在的涉及多种胁迫相应的可溶性蛋白,其含量非常丰富,占植物体内总可溶性蛋白的4%左右。自1975首次在马铃薯块茎中被发现以来,至今已发现其广泛存在于大麦、水稻、玉米、高粱、小麦、黑麦、绿豆、萝卜等60余种植物中,以本本科植物居多。nsLTPs蛋白含有8个半胱氨酸残(8CM;C-Xn-C-Xn-CC-Xn-CXC-Xn-C-Xn-C)、2个保守的五肽基序。由4个二硫键连接α-螺旋、连接环和C末端区域等构成稳定的三级结构,有热稳定性和化学稳定性,具有一个疏水性的空腔,可以转移多种类型的脂质小分子。
nsLTPs是一种多功能蛋白,参与植物多种生理过程,主要涉及植物生物膜间的磷脂运输、信号分子转导、脂质和角质的形成、表皮蜡质的生成、生殖与发育(细胞扩增、花粉形成、果实发育、种子萌发、根瘤形成)等。此外,越来越多研究发现,nsLTPs蛋白在植物免疫防御中也发挥着重要作用,包括响应植物生物胁迫防御和非生物胁迫防御。Fan等研究发现,过表达LTPs可增强欧洲油菜ROS清除酶活性,提高对核盘菌的抗性。Sarowar等研究发现,脂质转移蛋白基因LTP的过表达增强了烟草对植物病原菌的抗性,并在烟草系统的远距离信号传导中发挥作用。Zou等研究发现,玉米LTPs转导拟南芥,可以提高拟南芥的盐碱胁迫抗性。植物nsLTPs蛋白通过诱导免疫抑制病原菌生长,参与植物对各类病原菌的生物胁迫防御;通过信号传导、植物激素协作等方式,参与植物对ABA、干旱、盐碱、低温等非生物胁迫防御。因此,系统地分析植物nsLTPs蛋白结构、特性等生物特性,明确其在植物抗逆方面的作用,对于植物抗病抗逆研究及应用具有重要意义。
牡丹(Paeonia suffruticosa Andr.)属芍药科芍药属,为多年生木本落叶灌木。原产中国,有着有数千年的自然生长和1 600余年的人工栽培历史,是我国特有的观赏植物和药用植物物种,现引种遍布世界各地。近年来,随着牡丹籽油价值发掘,牡丹的用途又得到了进一步扩展,现已成为一种集观赏、药用、油用、食用、保健、工业用于一身的重要植物。近年来,国内外从种质资源调查、标准化栽培、组织培养、分子生物学、生理生化、丹皮酚提取、功能产品研发等方面对牡丹进行了系统性研究,但关于牡丹抗逆胁迫(抗寒、抗旱、抗盐碱、抗病虫等)方面的系统性研究较少,制约了牡丹抗逆育种方面的发展。笔者从NCBI GenBank數据库得到牡丹nsLTP基因cDNA序列,利用生物信息学在线软件对牡丹nsLTP基因及其他植物nsLTPs基因进行序列分析和功能预测,旨在为牡丹nsLTP家族基因系统性研究奠定基础,为牡丹抗逆方面功能基因研究提供新思路,也为其他植物nsLTPs基因的功能研究提供借鉴。
1 材料与方法
1.1 数据来源
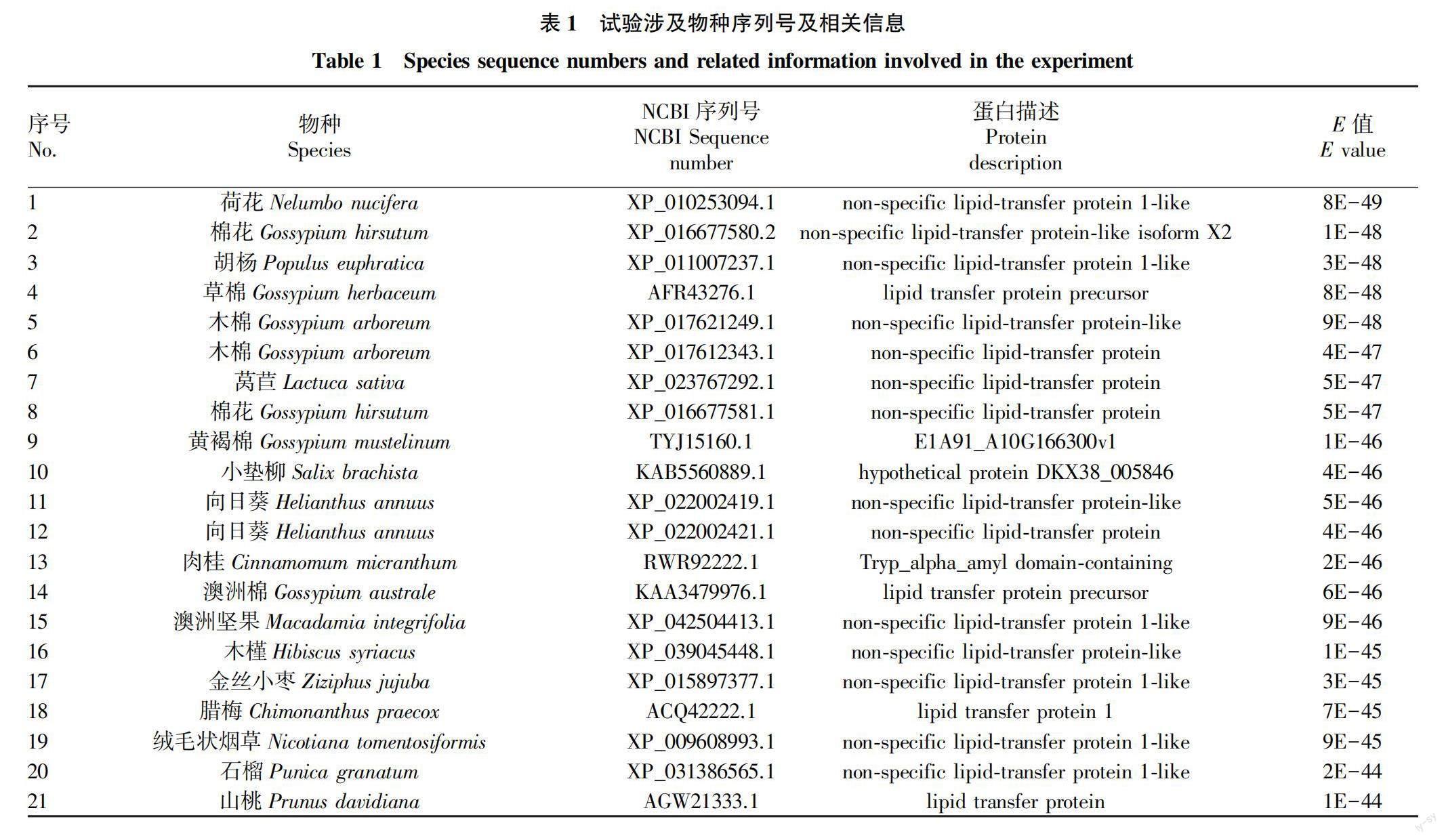
利用的牡丹nsLTP基因数据来源于美国国家生物技术信息中心( NCBI) 数据库,GenBank登录号为JZ840269.1。试验中所涉及的其他物种nsLTPs蛋白氨基酸序列均来源于NCBI数据库(表1)。
1.2 分析方法
利用NCBI-BLAST和NCBI-BLASTP从核苷酸及氨基酸水平分析牡丹nsLTP基因及其编码蛋白氨基酸序列特性。利用NCBI-ORF-Finder在线软件进行牡丹nsLTP开放阅读框ORF查找。EXpasy-ProtParam及SMS在线软件预测牡丹nsLTP蛋白的氨基酸组成、相对分子量、理论等电点、稳定系数等基本理化特性。利用ProtScale在线软件进行nsLTP蛋白亲疏水性分析。
用NetPhos 3.1 Server在线工具分析编码蛋白的潜在磷酸化位点。利用TargetP-2.0 Server及WOLF PSORT在线软件预测蛋白亚细胞定位信。利用SignalP-5.0 Server在线工具进行编码蛋白信号肽预测分析。利用TMHMM Server v.2.0进行编码蛋白跨膜结构域预测分析。NCBI BLASTP在线筛选同源蛋白序列,利用Bioedit软件进行多序列比对分析,用MEGA 6.0 软件N-J邻接法进行系统发育树分析。
利用SOPMA软件对PsLTP蛋白的二级结构进行预测分析。利用NCBI CDD在线预测编码蛋白的保守结构域。采用Swiss-model在线软件对牡丹nsLTP蛋白进行3D结构同源建模预测,Swiss-pdb viewer构建拉式构象图评估建模准确性。
2 结果与分析
2.1 牡丹PsLTP基因序列及蛋白氨基酸序列
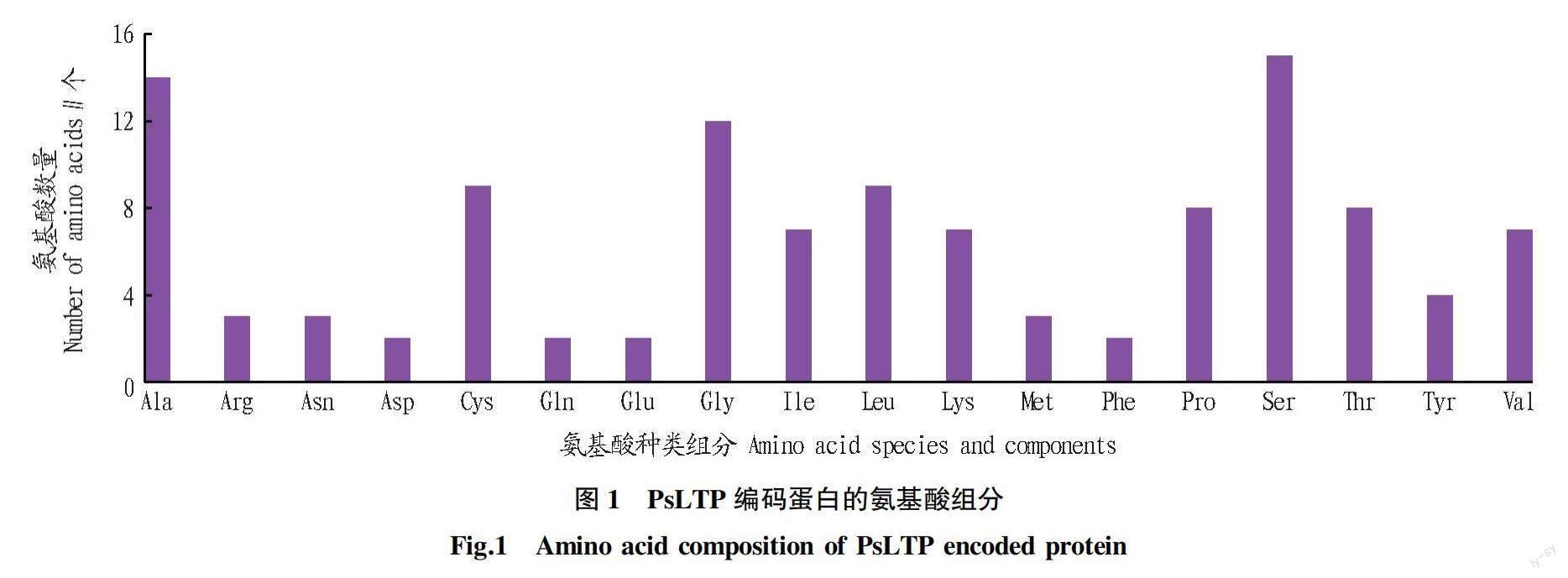
NCBI登陆的牡丹nsLTP基因序列cDNA长度为685 bp,SMS碱基分析显示,A有 191个(占比27.88%),T有172个(占比25.11%),G有163个(占比23.80%),C有159个(占比23.21%),GC含量占比为47.01%。ORF-Finder软件分析显示,牡丹nsLTP基因序列包含一段258 bp的3′-UTR,一段354 bp的开放阅读框(ORF)和一段77 bp的5′-UTR。开放阅读框(ORF)序列在第259~608个核苷酸区域蛋白编码117个氨基酸。EXpasy-ProtParam软件显示,该蛋白分子式C510H838N138O158S12,总原子数1 656个,相对分子量为11.815 kD,理论等电点为8.93。此外,该蛋白不稳定系数为42.67(大于40.00),属于不稳定性蛋白。由其氨基酸种类组分(图1)可知,天冬氨酸(Asp)、谷氨酰胺(Gln)、谷氨酸(Glu)及苯丙氨酸(Phe)数量最少,均为2个,组分占比1.71%;丝氨酸(Ser)数量最多15个,占比12.82%。
2.2 PsLTP蛋白亲/疏水性及磷酸化位点
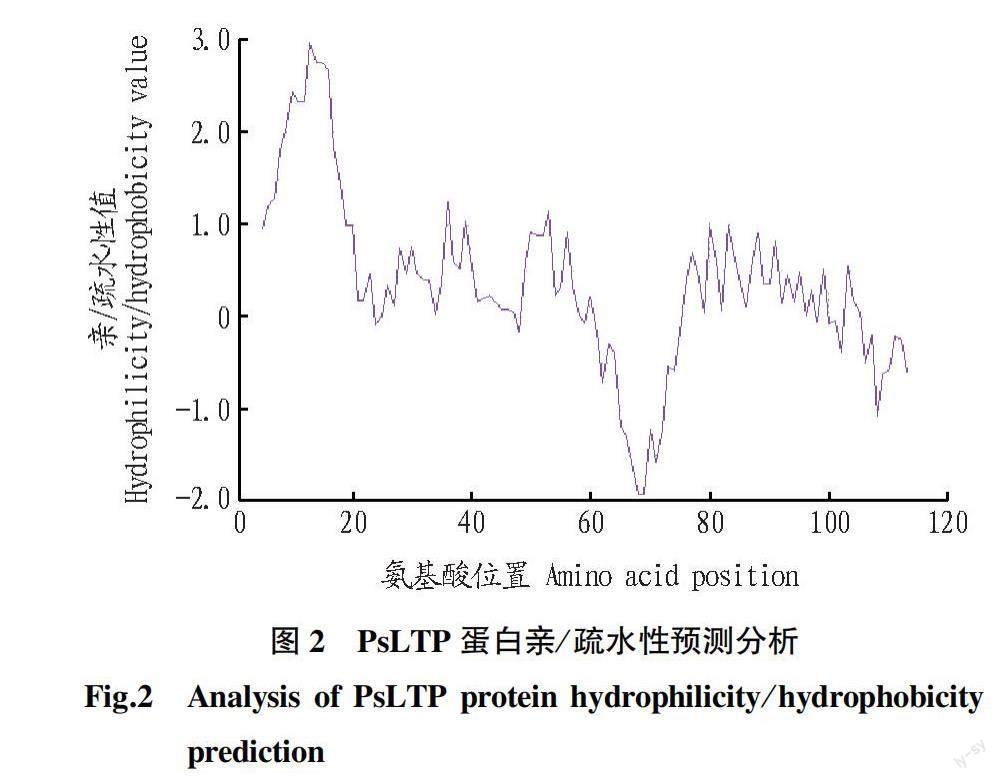
利用在线软件ProtScale对牡丹PsLTP蛋白进行亲/疏水性分析。负值表示亲水,正值表示疏水,绝对值越大,表示亲/疏水性越强。预测结果(图2)显示,牡丹PsLTP蛋白含有亲水性氨基酸30个,疏水性氨基酸79个。疏水性氨基酸数量占比高,总体平均亲/疏水性值为0.354,属于疏水性蛋白。牡丹PsLTP蛋白在Gly5~Tyr23氨基酸处具有1个明显的疏水性峰,在Asn61~Lys75位氨基酸处具有一个明显的親水性峰。其在第13位的丙氨酸Ala13处有最强疏水性峰值2.978,在第68位的脯氨酸Pro68处有最强的亲水性峰值-1.922。
利用在线软件NetPhos 3.1对牡丹PsLTP进行磷酸化位点分析。预测结果(表2)显示,PsLTP氨基酸序列包含了16个潜在的磷酸化位点(预测值大于0.5),其中8个位于丝氨酸(Ser,S),3个位于络氨酸(Tyr,Y),5个位于苏氨酸(Thr,T)。在整个氨基酸序列中,第82位氨基酸丝氨酸(Ser82)的磷酸化预测值最高为0.982,可能受磷酸化激酶激活。
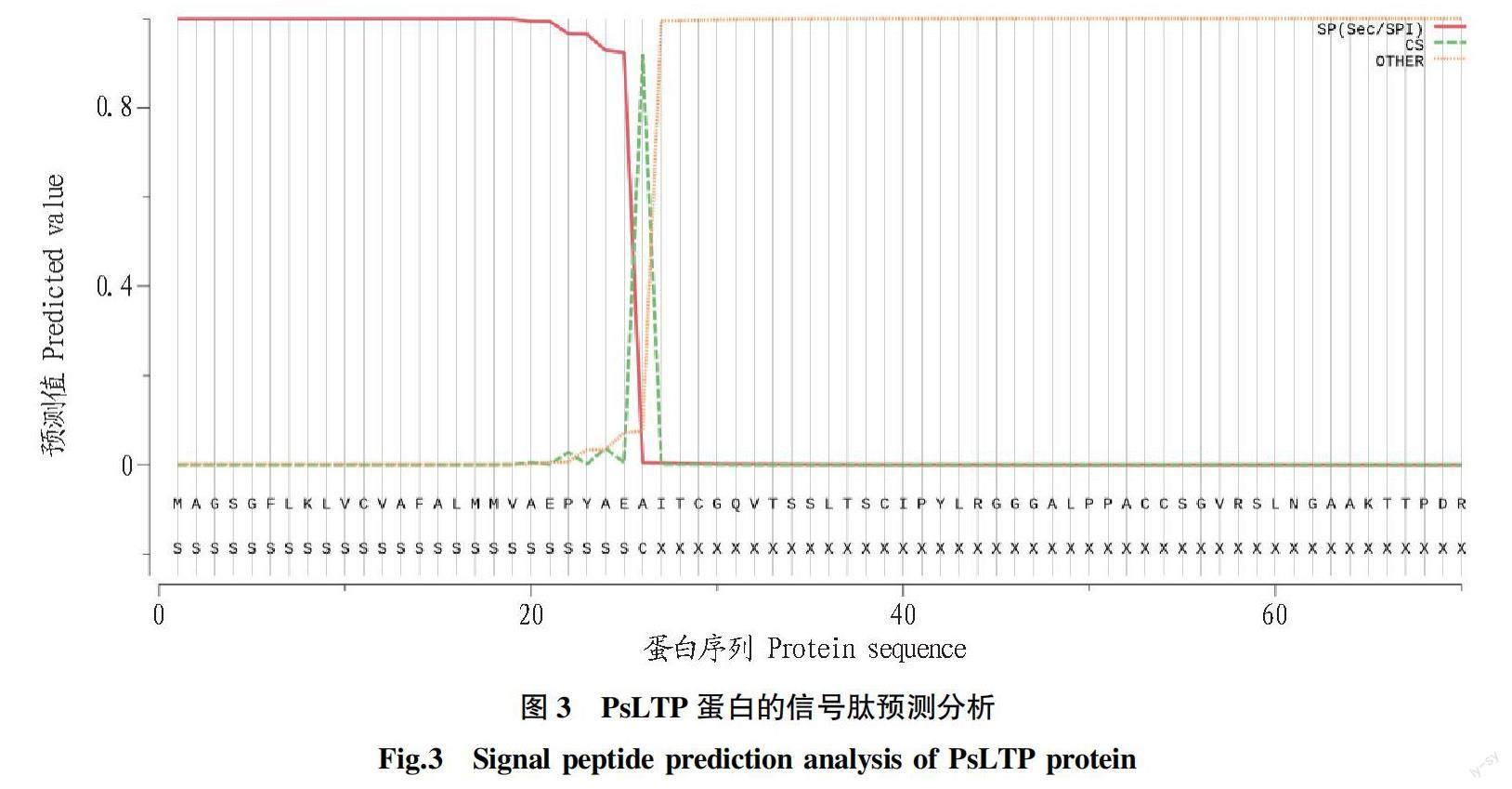
2.3 PsLTP蛋白信号肽和跨膜结构域
SignalP-5.0 Server在线软件对PsLTP蛋白进行信号肽分析,结果显示(图3):牡丹PsLTP蛋白分值曲线明显,有信号肽存在。信号肽位于第1~26位氨基酸残基之间,预测得分为0.920。信号肽与成熟肽的剪切位点介于第22和第23位氨基酸残基之间。TMHMM 2.0在线软件分析显示,PsLTP蛋白无跨膜结构域存在。
2.4 多序列比对及系统进化树
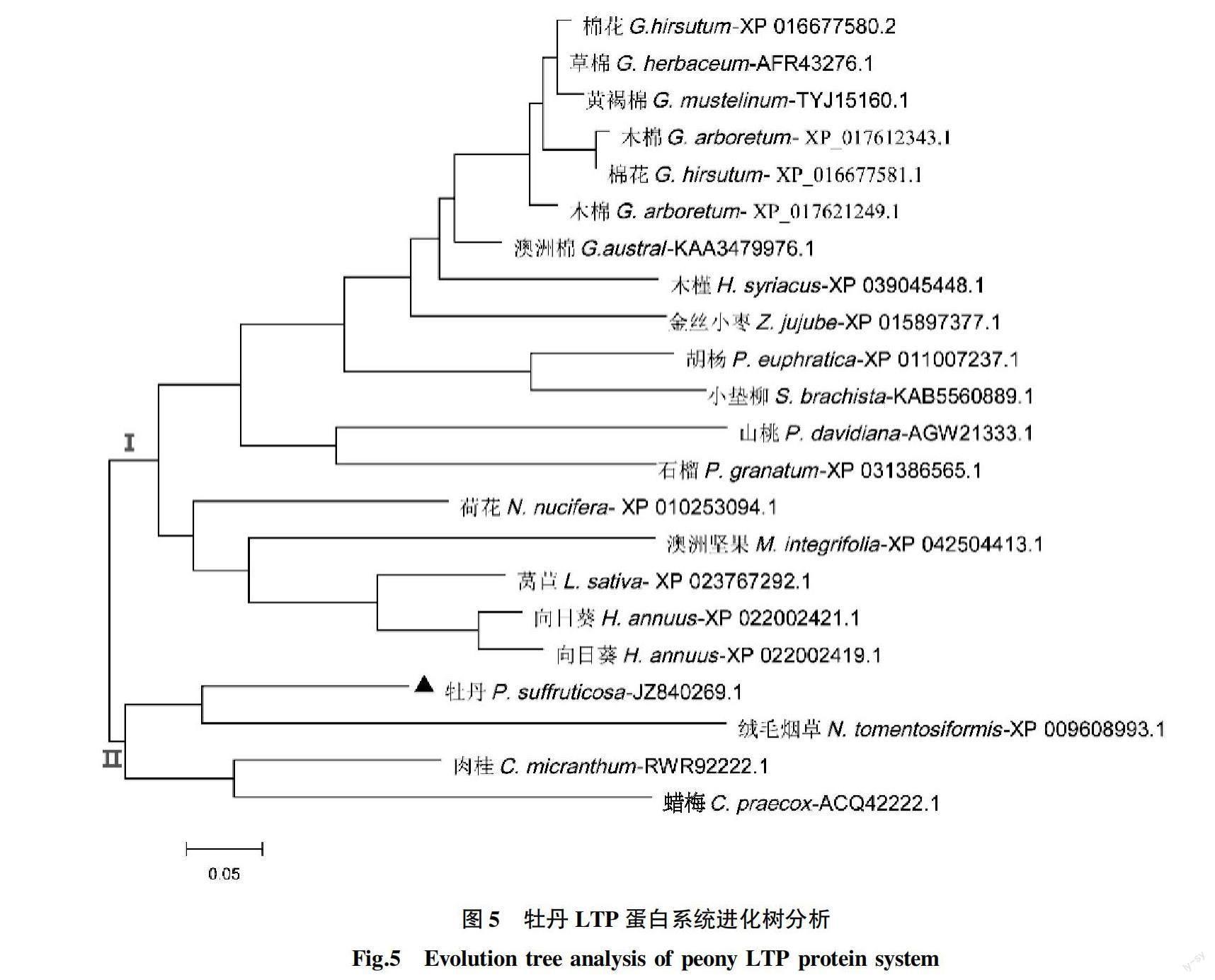
NCBI BLAST筛选得到21个nsLTPs的同源蛋白序列(表2),利用bioedit进行多序列比对分析(图4)。结果显示,多种植物nsLTPs蛋白序列的中间功能区域及Cys催化区域保守性高。牡丹nLTP蛋白(JZ840269.1)同荷花、棉花、胡杨、草棉、木棉等多种植物的nsLTPs蛋白具有同源性,与绒毛烟草XP_009608993.1同源性为62.7%。进一步通过MEGA软件进行多序列比对数据,以N-J法构建系统进化树(图4、5)。结果发现,22个植物LTPs蛋白氨基酸序列聚为2个大分支,棉花、草棉、木棉、木槿、向日葵等植物LTPs蛋白聚为分支 Ⅰ,牡丹、绒毛烟草、肉桂、蜡梅聚为分支Ⅱ。分支Ⅱ中,牡丹LTP蛋白(JZ840269.1)与绒毛烟草LTP蛋白(XP_009608993.1)聚为一小分支,肉桂LTP蛋白(RWR92222.1)与蜡梅LTP蛋白(ACQ42222.1)聚为一小分支。
2.5 保守结构域预测
利用NCBI 的BLASTP 在蛋白保守区数据库(conserved domain database)查找牡丹PsLTP蛋白的保守结构域,结果显示:牡丹PsLTP蛋白存在AAI-LTSS典型结构域(图6),该结构域具有α-淀粉酶抑制剂活性、脂类转移活性以及储藏蛋白功能,存在于诸多蛋白中,在植物抵御病虫害、跨膜运输脂类分子及植物种子营养物质贮藏等方面发挥了重要作用。
2.6 PsLTP蛋白高级结构的预测
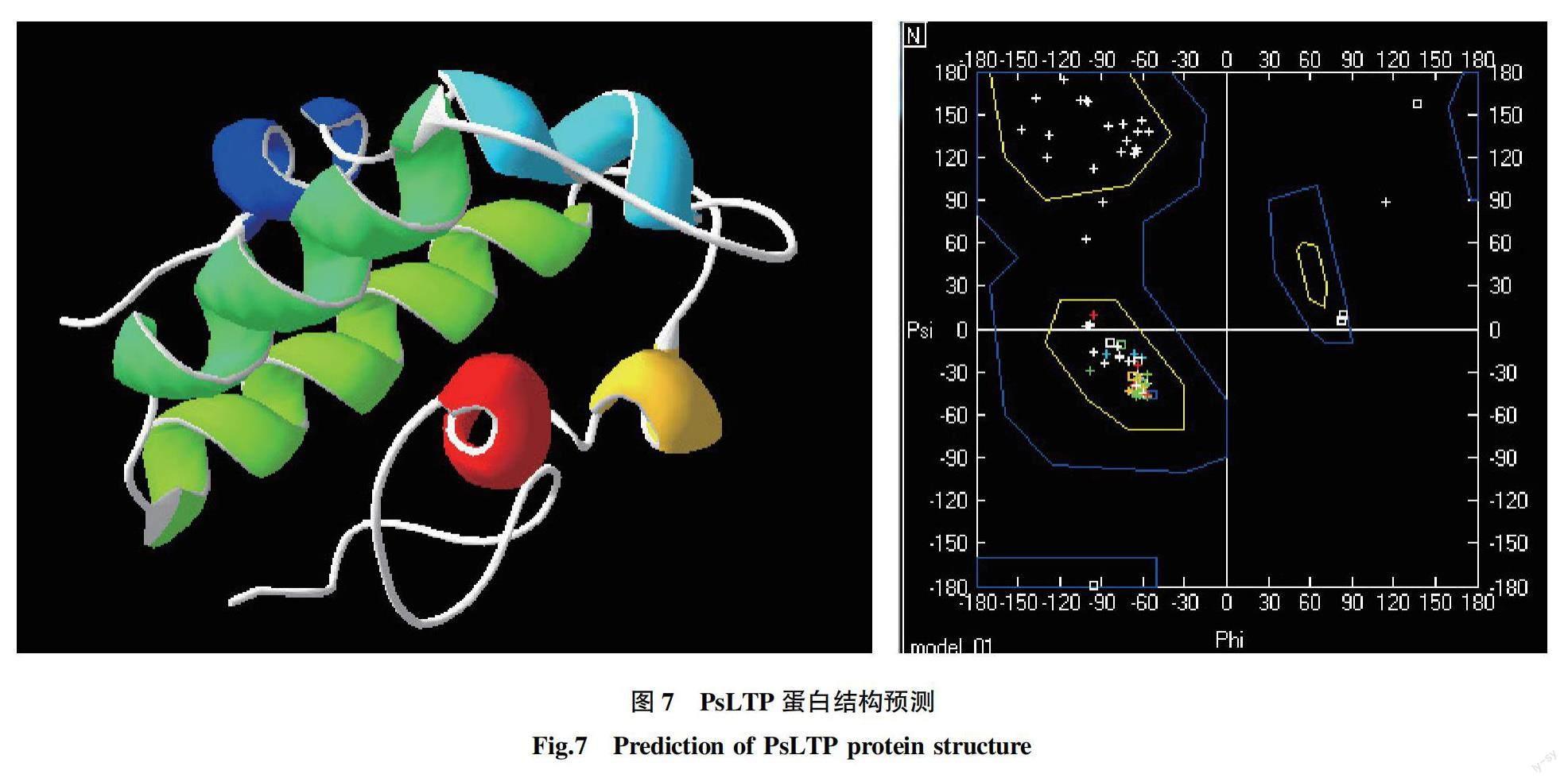
利用ExPaSy-SOPMA软件对PsLTP蛋白的二级结构进行预测(图7),结果显示,该蛋白的二级结构由45.3%的α-螺旋、43.59%的无规则卷曲、8.55%的延伸链和2.56%的β-转角组成。α-螺旋及无规则卷曲是该蛋白二级结构的主要成分,这些结构对其功能有重要作用。用SWISS-MODEL对PsLTP蛋白的三级结构进行预测分析,结果显示,PsLTP蛋白具有非特异性脂质转移酶蛋白的典型结构,Ramachan plots显示,牡丹PsLTP蛋白的三维结构Φ角和Ψ角位于黄色核心区域,其空间结构稳定,同源建模得到的三维结构可信度高。
3 结论与讨论
植物非特异性脂质转移蛋白是一类在高等植物中广泛存在的小分子碱性脂质结合蛋白,自从1975年首次在马铃薯块茎中分离出nsLTP,现已在水稻、豇豆、大麦、玉米等50余种植物分离出来。nsLTPs蛋白具有不同的生物学功能,在植物的很多生物进程中发挥着重要作用。如植物nsLTPs蛋白参与开花植物授粉、胚胎发育、脂质代谢、角质层合成、固氮作用、果实成熟、细胞凋亡、信号传导等。随着研究深入,更多研究发现,nsLTPs在植物免疫防御及抗逆方面也发挥重要作用。如部分植物nsLTPs蛋白对细菌、真菌、病毒及害虫具有抗性,参与了植物防御机制,部分植物nsLTPs蛋白通过高表达响应低温、干旱等非生物胁迫,参与植物抗逆。生物信息学是当今生命科学和自然科学的核心领域,是后基因组世代的重要研究方法。通过生物信息学系统分析牡丹nsLTP蛋白结构、特性等生物特性,明确其在植物抗逆方面的作用,旨在为牡丹nsLTP家族基因系统性研究奠定基础,为牡丹抗逆方面功能基因研究提供新思路。
该研究根据NCBI登陆的牡丹LTP基因序列,利用生物信息学分析该基因及编码蛋白的特性。基因序列分析显示,NCBI登陆的牡丹nsLTP基因序列cDNA长度为685 bp,GC含量占比47.01%。ORF显示,该蛋白编码117个氨基酸。蛋白氨基酸序列分析发现,分子式C510H838N138O158S12,蛋白中丝氨酸(Ser)数量最多15个,占比12.82%。蛋白不稳定系数为42.67,属于不稳定性蛋白。ProtScale预测蛋白含有亲水性氨基酸30个,疏水性氨基酸79个。疏水性氨基酸数量占比高,属于疏水性蛋白。NetPhos蛋白PsLTP氨基酸序列包含16个潜在的磷酸化位点。SignalP分析显示,牡丹PsLTP蛋白在第1~26位氨基酸残基之间有信号肽存在。多序列比对及系统进化树分析显示,牡丹LTP与绒毛烟草XP_009608993.1同源性为62.7%,聚在同一分支上,亲缘关系相对较近。蛋白结构预测分析显示,PsLTP蛋白具有非特异性脂质转移酶蛋白的典型结构,α-螺旋及无规则卷曲是该蛋白二级结构主要成分。LTP蛋白的序列及保守区结构的相对保守性为其他植物物种的LTP基因特性及结构功能研究奠定了基础。
该研究结果为深入研究牡丹LTP蛋白功能及抗逆防御机制提供了重要的基础信息,为牡丹抗逆方面功能基因研究提供新思路,也为其他植物nsLTPs基因的功能研究提供借鉴。通过对牡丹LTP的生物信息学分析,为今后植物LTP蛋白功能、载体构建、抗逆机制、抗病育种等工作的开展奠定了良好的研究与应用基础。
参考文献
[1]王海波,邹竹荣,龚明.小桐子非特异性脂质转移蛋白A的基因克隆及其表达和功能分析[J].植物生理學报,2015,51(1):127-135.
[2]THOMA S,KANEKO Y,SOMERVILLE C.A non-specific lipid transfer protein from Arabidopsis is a cell wall protein[J].Plant J,1993,3(3):427-436.
[3]SOSSOUNTZOV L,RUIZ-AVILA L,VIGNOLS F,et al.Spatial and temporal expression of a maize lipid transfer protein gene[J].Plant Cell,1991,3(9):923-933.
[4]CAMMUE B P A,THEVISSEN K,HENDRIKS M,et al.A potent antimicrobial protein from onion seeds showing sequence homology to plant lipid transfer proteins[J].Plant Physiol,1995,109(2):445-455.
[5]WANG S Y,WU J H,NG T B,et al.A non-specific lipid transfer protein with antifungal and antibacterial activities from the mung bean[J].Peptides,2004,25(8):1235-1242.
[6]WEI K F,ZHONG X J.Non-specific lipid transfer proteins in maize[J].BMC Plant Biol,2014,14:1-18.
[7]BOUTROT F,CHANTRET N,GAUTIER M F.Genome-wide analysis of the rice and Arabidopsis non-specific lipid transfer protein (nsLtp) gene families and identification of wheat nsLtp genes by EST data mining[J].BMC Genomics,2008,9:1-19.
[8]DUO J C,XIONG H Y,WU X X,et al.Genome-wide identification and expression profile under abiotic stress of the barley non-specific lipid transfer protein gene family and its Qingke Orthologues[J].BMC Genomics,2021,22(1):1-17.
[9]ODINTSOVA T I,SLEZINA M P,ISTOMINA E A,et al.Non-specific lipid transfer proteins in Triticum kiharae Dorof.et Migush.: Identification,characterization and expression profiling in response to pathogens and resistance inducers[J].Pathogens,2019,8(4):1-21.
[10]刘梅,生华,化文平,等.丹参非特异性脂质转移蛋白基因(SmLTP1)的克隆及其表达分析[J].植物生理学报,2011,47(1):63-68.
[11]DE OLIVEIRA,CARVALHO A,GOMES V M.Role of plant lipid transfer proteins in plant cell physiology-a concise review[J].Peptides,2007,28(5):1144-1153.
[12]LIU F,ZHANG X B,LU C M,et al.Non-specific lipid transfer proteins in plants:Presenting new advances and an integrated functional analysis[J].J Exp Bot,2015,66(19):5663-5681.
[13]SALMINEN T A,BLOMQVIST K,EDQVIST J.Lipid transfer proteins:Classification,nomenclature,structure,and function[J].Planta,2016,244(5):971-997.
[14]XU Y,ZHENG X X,SONG Y Z,et al.NtLTP4,a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum[J].Sci Rep,2018,8(1):1-14.
[15]FAN Y,DU K,GAO Y,et al.Transformation of LTP gene into Brassica napus to enhance its resistance to Sclerotinia sclerotiorum[J].Genetika,2013,49(4):439-447.
[16]SAROWAR S,KIM Y J,KIM K D,et al.Overexpression of lipid transfer protein (LTP) genes enhances resistance to plant pathogens and LTP functions in long-distance systemic signaling in tobacco[J].Plant Cell Rep,2009,28(3):419-427.
[17]ZOU H W,TIAN X H,MA G H,et al.Isolation and functional analysis of ZmLTP3,a homologue to Arabidopsis LTP3[J].Int J Mol Sci,2013,14(3):5025-5035.
[18]ZHANG J J,SHU Q Y,LIU Z A,et al.Two EST-derived marker systems for cultivar identification in tree peony[J].Plant Cell Rep,2012,31(2):299-310.
[19]ZHANG X P,XU Z D,YU X Y,et al.Identification of two novel R2R3-MYB transcription factors,PsMYB114L and PsMYB12L,related to anthocyanin biosynthesis in Paeonia suffruticosa[J].Int J Mol Sci,2019,20(5):1-18.
[20]駱俊,韩金蓉,王艳,等.高温胁迫下牡丹的抗逆生理响应[J].长江大学学报(自然科学版),2011,8(2):223-226.
[21]钟原,成仿云,吴静.牡丹(Paeonia sect.Moutan)功能基因研究进展[J].分子植物育种,2016,4(9):2353-2364.
[22]陈为,张琳.油用牡丹FAD2基因的生物信息学分析[J].北方园艺,2018(7):77-81.
[23]ZHANG L,LI D C,LIU L F.Paeonol:Pharmacological effects and mechanisms of action[J].Int Immunopharmacol,2019,72:413-421.
[24]冯德金,张雪莲,解志军.油用牡丹标准化种植技术[J].农村经济与科技,2018,29(7):67-68.
[25]彭东辉,张启翔,黄俊婷.中国野牡丹科观赏植物种质资源及其在福建省的分布初步调查[J].中国园林,2007,23(11):83-88.
