嘌呤信号在肝脏免疫调节中的作用
2023-07-05马发祥涂正坤
马发祥, 涂正坤
吉林大学第一医院肿瘤免疫实验室, 长春 130021
嘌呤在体内主要以嘌呤核苷酸的形式存在,除了具有关键的细胞内功能外, NAD+和ATP及其水解产物包括ADP、AMP和腺苷,是参与生理过程和病理状况的重要细胞外信号。这些嘌呤分子激活1型(P1)和2型(P2)两个嘌呤受体家族,从而激活不同的细胞内信号通路[1]。
肝脏由不同类型的细胞组成,包括肝细胞、肝星状细胞(HSC)、肝窦内皮细胞(LESC)、Kupffer细胞(KC)、肝窦内淋巴细胞等[1]。肝脏是体内代谢的主要器官,还具有重要的免疫功能,负责从体循环中清除病原体和外源性抗原。肝脏的关键部位及其独特的脉管系统使其能够从外周血中募集免疫细胞发挥固有免疫和适应性免疫的功能[2]。本文总结了肝脏环境中的嘌呤信号并讨论了其在慢性肝脏疾病中的作用。
1 嘌呤信号
嘌呤信号包括细胞内嘌呤释放到细胞外,以自分泌和旁分泌作用方式激活P1和P2嘌呤受体家族,以及通过胞外酶(CD38、CD39、CD73等)调节细胞外ATP/腺苷水平,以终止ATP核苷酸信号[1]。组织损伤引起释放的ATP对中性粒细胞、巨噬细胞、树突细胞和记忆T淋巴细胞具有趋化性并激活它们,NAD+也可以通过杀死某些T淋巴细胞来影响炎症。细胞外ATP和各种其他核苷酸激活P2X或P2Y受体以刺激炎症。随着时间的推移, ATP引起的炎症减弱甚至转为抗炎,这是由于调节细胞的募集和胞外酶(如CD39和CD73)的诱导,这些酶可将细胞外ATP快速转化为腺苷[3],腺苷通过免疫细胞上抗炎A2A和A2B腺苷受体发挥作用。
1.1 ATP和P2受体 ATP 被广泛认为是连接合成代谢和分解代谢的能量交换因子,并且还参与主动运输、运动收缩、磷酸化等。此外,细胞外ATP还被认为是一种重要的胞外信号,可通过细胞表面的P2受体发挥炎性作用。P2受体包括离子型P2X(P2X1~7)受体和代谢型P2Y(P2Y1、2、4、6、11~14)受体两个亚家族。P2X7受体已被广泛报道被ATP激活活化NLRP3炎性小体,活化的炎性小体通过capase1剪切IL-1β前体产生成熟的IL-1β并释放引起炎症[4]。已有研究[5-6]表明,ATP可以和P2Y2受体结合调节粒细胞和巨噬细胞的趋化性。
1.2 腺苷和P1受体 腺苷可通过经典途径和非经典途径分别由ATP和NAD+代谢产生,细胞外腺苷可通过P1受体发挥抗炎性作用。P1受体目前已被发现四种,即A1、A2A、A2B和A3腺苷受体。四种受体均为G蛋白偶联受体,A1和A3受体优先与Gi/o蛋白偶联,抑制腺苷酸环化酶和cAMP的产生,而A2A和A2B受体通常是触发细胞内cAMP积累的Gs偶联受体,引起抗炎反应的信号通路因细胞类型而异[7]。
1.3 腺苷产生经典途径的胞外酶 腺苷产生的经典途径由ATP代谢产生,首先由CD39水解ATP为AMP。CD39是一种跨膜蛋白,有两个跨膜结构域锚定在细胞膜上,并包含五个高度保守的序列结构域,被称为“腺苷三磷酸双磷酸酶保守区”,这些序列结构域参与活性位点形成和磷酸水解的细胞外核苷酸催化[8]。接着,CD73进一步水解AMP为腺苷。CD73是一种GPI锚定酶,包含一个N端结构域和一个C端结构域,它们由一个短α螺旋连接起来,N端结构域可以结合金属阳离子,而C端结构域结合AMP发挥催化作用,CD73也可以以可溶形式存在,具有与膜结合形式相似的活性[9]。
CD39和CD73的表达受到微环境中缺氧、氧化应激和炎性细胞因子的影响[10]。在转录水平上,CD39和CD73的表达被多个转录因子调控,如Sp1、HIF-1α、Stat3和Gfi-1[10-12]。CD39的启动子上存在转录因子Sp1的结合位点,Sp1在缺氧诱导的CD39表达中起关键作用[13]。缺氧还可通过HIF-1α结合CD73启动子增强CD73的表达[11]。在辅助性T淋巴细胞17(Th17)中,IL-6介导的Stat3转录因子激活和TGF-β驱动的Gfi-1转录抑制因子下调则可增加CD39和CD73的表达[12]。
1.4 腺苷产生非经典途径的胞外酶 腺苷也可由CD38、CD203a、CD73代谢NAD+产生。该途径绕过了CD39介导的经典途径,由CD38将NAD+转化为ADPR,ADPR随后被CD203a水解为AMP,AMP再被CD73水解为腺苷。在结构上,CD38 是一种具有单个跨膜片段的单链糖蛋白,由300个氨基酸组成[14]。CD203a是一种Ⅱ类跨膜糖蛋白,位于质膜和内质网中,以230~260 kD的同型二聚体形式存在,蛋白质的还原形式具有115~135 kD的分子大小,具体取决于细胞类型[15]。当微环境中不存在CD39的表达时,非经典途径可能作为一种主要的腺苷产生来源。
2 嘌呤信号在肝脏微环境中的作用
肝脏是人体最大的实质器官,在蛋白质、碳水化合物和脂类的代谢发挥关键功能,并通过胆汁清除有毒废物。同时,肝脏也被认为是以天然免疫为主的一个免疫特惠器官[16]。肝组织由实质性细胞(肝细胞)和非实质性细胞(NPC)组成。NPC主要包括LESC、KC、HSC和肝内免疫细胞。肝内免疫细胞包括树突细胞、自然杀伤(NK)细胞、NKT淋巴细胞、T淋巴细胞等。在正常生理情况下,从胃肠道经门静脉流向肝脏的血液中富含营养物质、细菌产物、毒素以及大量食物性抗原,而不引起肝脏炎症和损伤。在病理情况下,大多数病原体在肝脏成功清除,甚至一些已建立感染的病原体最终也会被肝脏免疫反应所清除。这一过程中,肝脏组织独特的微环境和肝窦内免疫细胞包括肝细胞、NPC、 KC、NK细胞和T淋巴细胞调节肝脏区域免疫反应,从而决定肝脏免疫和耐受[16]。肝脏微环境中复杂的免疫调节机制目前尚不完全清楚。
嘌呤信号包括ATP信号和腺苷信号,受到胞外酶和嘌呤受体在肝脏驻留细胞及肝脏募集的免疫细胞表面表达调控。生理状态下,ATP的浓度和腺苷浓度被胞外酶控制在稳定的平衡状态以维持肝脏的正常代谢功能。肝脏疾病会导致嘌呤信号的失衡,影响肝脏内各种细胞的功能(图1)。
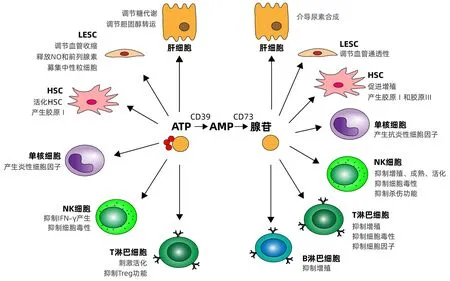
注:Treg,调节性T淋巴细胞。图1 嘌呤信号对肝脏中细胞的调节Figure 1 Regulation of cells in the liver by purine signaling
2.1 肝细胞 肝细胞是肝脏中主要的实质细胞,约占肝脏质量的80%。肝细胞执行肝脏的主要代谢活动。肝细胞上CD73表达与Mallory-Denk 小体的形成相关进而导致对肝病的易感性[17]。肝细胞表面P2X4受体激活调节细胞体积[18],P2X7受体激活介导肝细胞Ca2+依赖性的杀伤作用[19]。P2Y受体和P1受体的激活多与肝细胞的代谢功能相关, P2Y1和P2Y2受体激活调节糖原代谢[20-21],P2Y13受体激活调节胆固醇转运[22],P1受体激活主要与尿素合成相关[23]。
2.2 LESC LSEC形成可渗透屏障,充当血细胞、HSC和肝细胞之间的界面,具有非常高的内吞能力。LESC上有CD39和CD73的表达,部分肝切除术后,CD39 的表达在LESC上增加并且与肝再生相关[24]。此外,由LESC上CD39和CD73产生的腺苷可通过减少中性粒细胞积累限制炎症[25]。P2Y受体与血管收缩相关,介导LESC释放NO和分泌前列腺素,P2X7激活会募集中性粒细胞引发炎性反应[26]。缺氧状态下,A2B受体上调,它的激活可调节血管的通透性变化。
2.3 HSC HSC具有两种表型:静止的维生素 A 储存细胞和活化的肌成纤维细胞样细胞,在肝损伤的情况下负责胶原沉积。HSC在静止和激活条件下均表达 CD39 mRNA[27]。在肌成纤维细胞上CD73高表达而CD39并未观察到表达[28]。静止的HSC表达P2Y 亚型 P2Y2 和 P2Y4,激活的HSC表达 P2Y 亚型 P2Y6,P2Y 受体将细胞外 ATP 与肌醇三磷酸介导的细胞溶质钙信号联系起来,从而导致局部钙信号传导和细胞收缩[28]。HSC上A2A受体的激活抑制HSC收缩功能[29],增强增殖功能[30],还可以增加Ⅰ型胶原蛋白和Ⅲ型胶原蛋白的生成[31-32]。
2.4 单核/巨噬细胞 肝脏中的单核/巨噬细胞包括从外周血募集的单核巨噬细胞和肝脏驻留的KC。KC是人体最大的单核/巨噬细胞群,占肝脏中的NPC约35%和所有组织巨噬细胞的90%。单核细胞表达CD39但很少表达CD73,巨噬细胞CD73和CD39都有表达。巨噬细胞表达的CD39通过将ATP水解最终产生腺苷减弱P2X7引起的炎症并减弱脓毒症引起的肝损伤[33]。在感染和肿瘤中也发现了单核/巨噬细胞胞外酶的上调导致ATP被水解产生腺苷,促进感染和肿瘤进展[34-35]。单核/巨噬细胞表达P2X7受体,其激活会活化NLRP3炎性小体导致IL-1β的大量释放引起炎症[36]。巨噬细胞表面表达A2A和A2B受体,腺苷可激活A2A和A2B受体产生抗炎性细胞因子IL-10抑制炎症[34]。
2.5 NK细胞 肝脏含有体内数量最多的NK细胞。肝脏内所有淋巴细胞的三分之一到二分之一可以定义为NK细胞。通常这些先天性淋巴细胞在大多数其他组织中很少见,但在筛查感染和组织病理学方面发挥着关键作用。健康个体的NK 细胞中很少有CD73 的表达,但是在受到环境影响时,CD73可以在NK细胞上表达。已经有报道在肿瘤微环境和暴露于间充质干细胞时,NK细胞CD73的表达被显著上调[37-38]。CD39也已经被报道在NK细胞上表达并且CD39缺失调节IFN-γ的分泌减轻小鼠的肝缺血/再灌注损伤[39]。在肿瘤微环境中,CD73和CD39表达导致的腺苷产生抑制NK细胞反应进而导致免疫抑制,CD39的抑制剂可作为一种药物恢复NK细胞的细胞毒性[40]。P2X和P2Y受体在NK细胞上表达,它们的激活会抑制 NK 细胞分泌 IFN-γ,抑制 NK 细胞毒性,促进肝再生[41]。A2AR是NK细胞上主要表达的腺苷受体,激活抑制 NK 细胞成熟、增殖、活化、细胞毒性细胞因子(如IFN-γ和TNF-α)的产生,以及对靶细胞的杀伤功能[42-45]。
2.6 T淋巴细胞 T淋巴细胞是淋巴细胞的主要组分,是细胞免疫的主要效应细胞。在肝脏内,常规T淋巴细胞包括 CD8+和 CD4+T淋巴细胞,这些受体分别在 MHC Ⅰ类和Ⅱ类分子的背景下识别抗原。肝脏中的CD8+T淋巴细胞的数量通常超过 CD4+T淋巴细胞并且效应细胞/记忆细胞的频率高于血液中的频率。CD39在Treg上表达,CD73在小鼠的Treg上与CD39共表达,但是在人类的Treg中仅有很少一部分表达CD73[9]。除Treg外, CD8+T淋巴细胞和Th17也表达CD73和CD39,并抑制免疫反应[46-47]。P2X7受体在各种T淋巴细胞表面均有表达,被激活后抑制Treg的功能并阻断产生IL-10的CD8+T淋巴细胞发育[46]。腺苷受体A2AR 在效应T淋巴细胞的炎症过程中上调[48]。其激活抑制效应T淋巴细胞增殖[49]、细胞毒活性和细胞因子产生(如TNF-α、IFN-γ、IL-2),导致外周免疫抑制。
2.7 B淋巴细胞 B淋巴细胞是体液免疫应答的主要细胞,通过产生抗体、抗原递呈和产生细胞因子的方式发挥免疫功能。通常,B淋巴细胞活化、分化和增殖发生在次级淋巴器官的淋巴滤泡中,肝内生发中心IgM、IgD、IgG阳性B淋巴细胞分布及 Ki-67、CD23 或 bcl-2、bcl-6 基因产物的表达模式与淋巴结相似,提示肝内生发中心作为功能性滤泡结构发挥作用。人类B淋巴细胞对于CD39和CD73往往是共表达[50]。B淋巴细胞上CD39和CD73通过产生腺苷发挥免疫抑制的功能。B淋巴细胞表达CD39和CD73产生腺苷可通过抑制巨噬细胞的杀菌功能和抑制T淋巴细胞增殖等方式抑制免疫应答[50-51]。除直接表达胞外酶外,B淋巴细胞还可以释放表达CD39和CD73的囊泡抑制其他细胞的免疫功能[52]。B淋巴细胞上P2受体的表达和功能目前并没有明确报道。Saze等[50]发现B淋巴细胞表达A1、A2A和A3受体,但不表达A2B受体,并且A3受体的激活抑制B淋巴细胞增殖。
3 嘌呤信号在肝脏疾病中的作用
3.1 肝炎
3.1.1 ATP通过P2受体引发炎症 有氧条件下线粒体氧化磷酸化是肝细胞ATP产生的主要来源[53]。生理状态下的胞外ATP水平受到严格的控制,通常维持在微摩水平,控制ATP释放的机制包括分泌性胞吐作用、连接蛋白或 pannexin半通道、ATP 结合盒 (ABC) 转运蛋白等[10]。然而在肝脏炎症发生时,往往会导致肝细胞的损伤,ATP会从损伤的细胞中释放出来,细胞外的ATP浓度显著增加。ATP是P2受体的配体,P2 受体家族由 P2Y G蛋白偶联受体和 P2X (P2X1~7) 配体门控离子通道组成。P2X7 受体已被广泛研究,细胞外的ATP激活特异性P2X7受体活化NLRP3炎性小体,活化的炎性小体剪切IL-1β前体,进而导致IL-1β的大量释放[54]。在伴刀豆球蛋白 A (Con A) 诱导的急性肝炎中,大量 ATP 从肝脏释放并诱导 P2Y2受体表达,增强细胞炎性反应并促进肝细胞死亡[55]。此外还有研究[56]表明,ATP除了可以直接增强免疫细胞炎性反应外,还可以通过P2Y受体激活使炎症细胞向组织损伤部位募集,发挥炎症效应。
3.1.2 腺苷通过P1受体限制炎症 ATP可通过胞外酶代谢为腺苷。为了限制炎症,肝脏募集表达高水平 CD39 和 CD73 的免疫细胞,这些免疫细胞一方面可以清除ATP减弱炎症反应,另一方面可以产生免疫抑制性的腺苷发挥抗炎功能。Savio等[33]的研究证明了巨噬细胞中的 CD39清除 eATP 并最终产生腺苷来减轻败血症相关的肝损伤。腺苷对免疫细胞P1受体的激活可直接发挥抗炎作用。巨噬细胞是产生炎性细胞因子和抗炎性细胞因子的主要细胞,腺苷可抑制巨噬细胞TNF-α、IL-12的产生,增加IL-10的产生[34,57],但对于影响细胞因子产生的机制根据研究所选择的细胞不同其发挥作用的受体也不同,主要是A2A和A2B受体发挥作用。对于NK细胞,腺苷抑制 NK 细胞的细胞毒活性和细胞因子产生[44]。关于T淋巴细胞的研究中发现,A2A是腺苷发挥作用的主要受体,A2A受体激活可抑制幼稚 CD4+T淋巴细胞分泌 IL-2[58],从而减少其在T淋巴细胞受体刺激后的增殖,A2A受体激活还抑制初始 CD4+T淋巴细胞和极化 Th1、Th2 细胞产生的 IFN-γ及IL-4[59]。除免疫细胞外,腺苷也可以通过A2B作用于内皮细胞调节血管的屏障功能防止免疫细胞在组织中聚集而减弱炎症反应[60]。病毒性肝炎是常见的慢性肝病之一,其特点是免疫抑制导致病毒持续感染。研究[61]发现HBV感染者中有更高比例的CD39+Treg并与疾病进展相关,此外在戊型肝炎患者的外周血中检测到 Treg 上 CD73 的表达显著升高[62],说明腺苷导致的免疫抑制可能是病毒性肝炎中病毒持续存在的原因。
3.2 肝纤维化
3.2.1 嘌呤信号作用于HSC导致肝纤维化 多种原因引起的肝损伤使肝形成瘢痕组织导致肝纤维化。如果瘢痕形成过程持续一段时间,纤维化就会变成永久性的发展为肝硬化。引起肝纤维化的主要细胞成分是HSC,它分化为肌成纤维细胞而导致胞外基质的积累。静止的HSC表达P2Y2R和P2Y4R,而活化的HSC表达P2Y6R,这些受体均可被ATP和UTP激活,HSC中 P2Y受体的激活使前胶原Ⅰ转录增强三倍,说明P2Y受体可能在纤维化中有关键作用[27]。有研究[63]表明,P2X7 受体可以通过 PKC 依赖性 GSK3β 通路介导乙醛诱导的HSC活化升高胶原的蛋白质和 mRNA水平,抑制P2X7R可减轻小鼠模型中的肝损伤和纤维化。Le Guilcher等[64]研究表明,当敲除P2X4R基因或对P2X4R给予药理学抑制可保护小鼠免于肝纤维化和肝肌成纤维细胞积累。在小鼠的肝纤维化标本中发现肝细胞间的纤维化隔膜和肝窦间隙中CD73表达增加[28],敲除CD73后小鼠肝脏释放的腺苷水平降低,肝脏中的胶原蛋白含量也明显降低从而防止肝纤维化[65]。关于P1受体在纤维化中作用的报道主要集中在A2AR上,TAA诱导的肝纤维化小鼠肝脏A2AR表达增加[32],腺苷激活A2AR可促进HSC的增殖和Ⅲ型胶原的表达[30,32]。A2AR缺陷小鼠可防止小鼠发生纤维化[32],A2AR 拮抗剂治疗既可以预防也可以逆转乙醇诱导的肝纤维化[66]。以上这些研究表明无论是ATP信号还是腺苷信号都在肝纤维化中发挥着促纤维化作用,P1和P2受体可作为纤维化治疗的潜在靶点。
3.2.2 嘌呤信号通过调节炎性反应调节肝纤维化 炎症在纤维化过程的任何阶段,甚至在纤维化消退过程中都起着关键作用。在肝脏炎症期间,免疫细胞释放大量炎性和抗炎性细胞因子,这些细胞因子对于HSC的激活具有调节作用。ATP和腺苷分别作用于免疫细胞调节炎性和抗炎性细胞因子的产生。TNF-α和IL-1是ATP炎性信号中产生的两种主要细胞因子。在肝纤维化过程中,TNF-α在HSC的活化和细胞外基质的合成中起重要作用[67],IL-1可直接激活HSC,刺激其产生MMP-9、MMP-13和TIMP-1,导致肝纤维化[68]。腺苷可诱导免疫细胞产生TGF-β和IL-10。TGF-β 是已知最强的肝纤维化诱导剂,TGF-β1已被证明在肝纤维化的启动和维持中发挥关键作用,TGF-β1可通过多方面作用促纤维化,包括刺激HCS活化,诱导基质产生基因表达,抑制细胞外基质降解等[69-70]。IL-10可下调促炎反应并对肝纤维化具有调节作用,IL-10 已被证明通过抑制 HSC 活性发挥抗纤维化作用[70],IL-10对于肝纤维化患者可能具有治疗潜力。因此腺苷通过免疫细胞对纤维化的作用可能是双向的。
3.3 肝细胞癌(HCC) 慢性肝炎和肝硬化最终可发展为HCC。从坏死细胞中释放的ATP 作为免疫系统的早期危险信号在肿瘤进展过程中起着重要作用。与健康肝组织相比,肝肿瘤组织中P2受体的表达增加[71-72]。P2X3 嘌呤能受体高表达与无复发生存率差相关,而 P2Y13 高表达与无复发生存率改善相关,细胞外ATP 可激活P2X3 受体,促进 HCC细胞的增殖[71]。P2Y11R在人类 HCC 中特异性表达促进ATP 诱导的 Ca2+信号传导和细胞迁移。P2Y2信号也可以通过ATP介导的Ca2+信号传导促进HCC细胞迁移。在肝脏肿瘤环境中有大量胞外酶表达,通过产生腺苷促进肿瘤进展。最近的研究[35]发现HCC上表达的CD73和巨噬细胞上表达的CD39协同作用产生腺苷抑制T淋巴细胞功能。P2受体在HCC上和各类免疫细胞上都有表达,腺苷可通过HCC表面A2AR激活PI3K/AKT 信号促进肿瘤增殖和转移[73],免疫细胞上A2AR的激活则会抑制免疫应答促进肿瘤逃逸。
4 小结
ATP和腺苷在胞外可作为不同的两种信号发挥炎性和抗炎性的作用,胞外酶可以将ATP水解为腺苷(图2)。肝脏微环境中的肝脏驻留细胞和肝脏募集的免疫细胞表面有P1受体、P2受体和胞外酶的表达。
肝脏炎症发生时,肝脏通过调节胞外酶的表达减少ATP引发的炎性反应,增加腺苷的抗炎性反应来限制炎症。嘌呤信号可通过直接作用的方式和调节炎性及抗炎性反应影响肝脏疾病进展。
利益冲突声明:本文不存在任何利益冲突。
作者贡献声明:马发祥负责查阅文献,撰写文章;涂正坤负责拟定文章思路,指导撰写文章并最后定稿。
