采后贮藏时间对不同发育期采收的巨大口蘑子实体褐变及相关酶活性的影响研究
2023-04-14宋舒晴吴英韩颖峰郑晓欣刘欣芊莫美华
宋舒晴 吴英 韩颖峰 郑晓欣 刘欣芊 莫美华
(华南农业大学食品学院,广州 510642)
巨大口蘑[Macrocybe gigantea(Massee)Pegler& Lodge],又名大白口蘑,是一种丛生的高温型珍稀食用菌,主要分布在热带及亚热带地区[1]。巨大口蘑子实体肥大,菌肉鲜美,营养价值高(多糖、粗蛋白和矿质元素含量较高,氨基酸种类齐全)[2-3],且具有抗肿瘤、抗氧化、抑制高血压和抑制细菌、真菌、艾滋病毒等作用[4-7],故其市场需求量较大。
研究与实践均表明,食用菌褐变是降低其品质的重要因素之一,其中酶促褐变是其褐变的主要原因。酶促褐变主要是指褐变相关酶和酚类底物在存在氧气的情况下。生成醌类物质,并进一步发生反应促进醌类物质生成,进而产生黑色素并导致食用菌表面褐变;与酶促褐变相关的酶类主要有多酚氧化酶、过氧化物酶、过氧化氢酶、苯丙氨酸解氨酶、脂肪氧合酶等[8-10]。有研究表明,巨大口蘑子实体在8~12 ℃下可保存30 d 不变色、不腐烂,耐储性很好,故研究其不易褐变的机理对于食用菌的贮藏及保鲜具有重要意义[11-13]。在此背景下,笔者于2021年对不同发育期采收的巨大口蘑子实体在采收后不同贮藏期的褐变相关生理指标进行了测定,以期探明巨大口蘑子实体在贮藏过程中生理生化指标的变化规律。
1 材料与方法
1.1 试验材料
巨大口蘑SCAU1菌株由华南农业大学应用真菌研究室提供,栽培于启林北食用菌基地。
1.2 主要试剂与仪器
主要试剂为:无水乙醇(购于国药集团化学试剂公司),磷酸二氢钠、磷酸氢二钠、醋酸钠、甲醇、盐酸(购于广州化学试剂厂),邻苯二酚(购于天津市福晨化学试剂厂),邻联甲苯胺(购于上海晶体试剂有限公司),冰醋酸(购于广东东华化学试剂厂),L-苯丙氨酸(购于Solarbio 公司)。本试验所用试剂均为分析纯。
主要仪器为:PB-10 型pH 计[赛多利斯科学仪器(北京)有限公司生产],UV-1100 型紫外可见分光光度计(上海美谱达仪器有限公司生产),X-RITE SP62 型色差仪(美国爱色丽公司生产),GL-16GC 型高速冷冻离心机(上海安亭科学仪器厂生产)。
1.3 试验方法
1.3.1 样品处理
依据巨大口蘑子实体发育标准[14-17],将其分为原基期、菇蕾期、幼菇期、中菇期、半球期、平展期、卷边期、衰老期等8个不同的发育期,然后分别在8个发育期采收无病虫害、无损伤、符合试验要求的子实体,各50 g,重复3 次,用PE 气调保鲜袋封口包装,在温度为12 ℃、相对湿度为95%的生化培养箱中进行避光贮藏。
1.3.2 褐变指数测定
参照杨春敏等[18]的方法,取1.00 g巨大口蘑子实体,使用浓度为95%的乙醇5.00 mL,在冰浴条件下将其研磨成浆,测定其OD420吸光值(吸光值越大,子实体褐变程度越大),各处理褐变指数测定重复3 次。
1.3.3 多酚氧化酶活性测定
参照王治江等[19]的方法,测定多酚氧化酶活性OD4205 min 内的变化,酶活力单位以每分钟变化0.001 表示,单位为U/gFW,各处理多酚氧化酶活性测定重复3 次。
1.3.4 酪氨酸酶活性测定
参照魏要武等[20]的方法,测定酪氨酸酶活性OD4805 min 内的变化,酶活力单位以每分钟变化0.001 表示,单位为U/gFW,各处理酪氨酸酶活性测定重复3 次。
1.3.5 漆酶活性测定
参照潘迎捷等[21]的方法,测定漆酶活性OD6005 min 内的变化,酶活力单位以每分钟变化0.001 表示,单位为U/gFW,各处理漆酶活性测定重复3 次。
1.3.6 苯丙氨酸解氨酶活性测定
参照吴英等[22]的方法,在OD290nm 处检测吸光值,以每分钟OD290减少0.001 为1 个酶活力单位,单位为U/g F W,各处理苯丙氨酸解氨酶活性测定重复3 次。
1.3.7 总酚含量测定
参考刘丽丹[23]的方法进行总酚含量测定,单位为mg/mL,各处理总酚含量测定重复3 次。其中,以焦性没食子酸为标准品,建立总酚含量的标准曲线,见图1。

图1 总酚含量的标准曲线
1.4 数据统计
试验数据均采用SPSS 24.0 软件进行统计和方差分析,采用Duncan’s 新复极差法进行差异显著性分析,采用Excel 2007 软件作图。
2 结果与分析
2.1 不同发育期采收的巨大口蘑子实体的褐变度变化情况
由表1可知,不同发育期采收的巨大口蘑子实体的褐变度,均随贮藏时间的延长大体上呈缓慢上升的趋势。贮藏15 d 内,不同发育期采收的巨大口蘑子实体的褐变度变化相对较平缓,这体现出巨大口蘑子实体不易褐变的特性。贮藏18 d 时,原基期、卷边期、衰老期采收的巨大口蘑子实体的褐变度分别为0.551、0.226、0.317,而幼菇期、中菇期、半球期、平展期采收的巨大口蘑子实体的褐变度分别为0.148、0.050、0.197、0.111。以上结果表明,原基期、卷边期、衰老期采收的巨大口蘑子实体比幼菇期、中菇期、半球期、平展期采收的巨大口蘑子实体更易发生褐变。考虑到经济效益问题,巨大口蘑子实体应尽量选择在中菇期、半球期和平展期进行采收。

表1 不同发育期采收的巨大口蘑子实体的褐变度变化情况
2.2 不同发育期采收的巨大口蘑子实体的多酚氧化酶活性变化情况
由表2可知,不同发育期采收的巨大口蘑子实体的多酚氧化酶活性,均随贮藏时间的延长大体上呈波浪式下降的趋势。刚采摘时,原基期采收的巨大口蘑子实体的多酚氧化酶活性显著高于其他发育期采收的子实体(p<0.05);贮藏15 d 内,不同发育期采收的巨大口蘑子实体的多酚氧化酶活性下降较快;贮藏15 d 后,不同发育期采收的巨大口蘑子实体的多酚氧化酶活性下降趋于平缓;贮藏30 d时,不同发育期采收的巨大口蘑子实体的多酚氧化酶活性均降至最低,且处理间多酚氧化酶活性差异不显著(p<0.05)。经分析,巨大口蘑子实体的多酚氧化酶酶活性低且呈现下降的趋势,这与巨大口蘑子实体不易褐变密切相关。同时,在整个贮藏过程中,原基期、卷边期、衰老期采收的巨大口蘑子实体的多酚氧化酶活性相对较高,菇蕾期、幼菇期、中菇期、半球期、平展期采收的巨大口蘑子实体的多酚氧化酶活性相对较低,该结果与原基期、卷边期、衰老期采收的巨大口蘑子实体的褐变度高,而其他发育期采收的巨大口蘑子实体的褐变度较低这一结果相吻合。
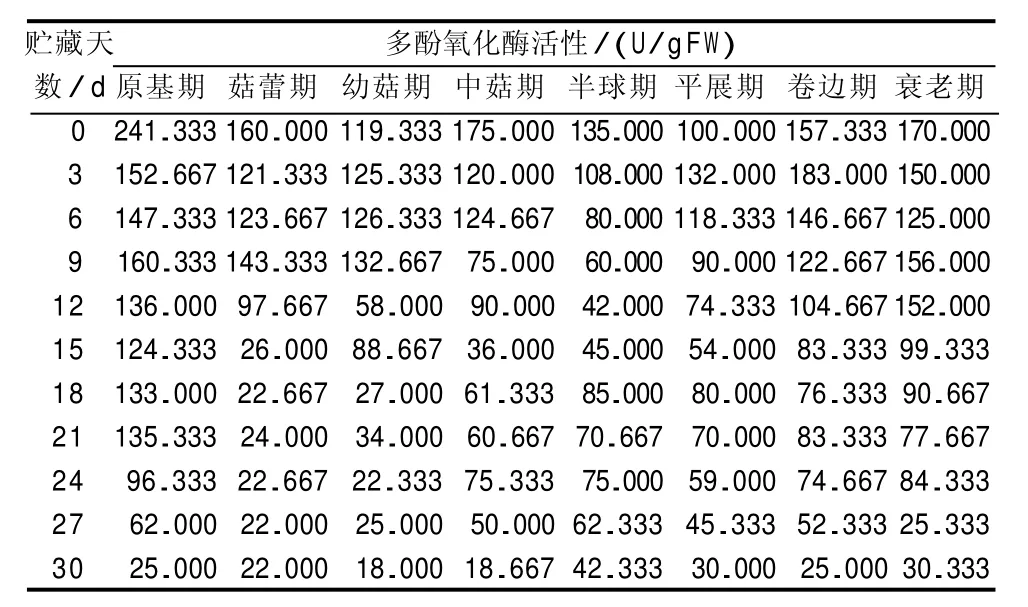
表2 不同发育期采收的巨大口蘑子实体的多酚氧化酶活性变化情况
2.3 不同发育期采收的巨大口蘑子实体的酪氨酸酶活性变化情况
由表3可知,不同发育期采收的巨大口蘑子实体的酪氨酸酶活性,均随贮藏时间的延长在一定范围内呈波浪式变化的趋势,且保持相对恒定。贮藏3 d时,衰老期采收的巨大口蘑子实体的酪氨酸酶活性出现第1 个峰值,而其他发育期采收的巨大口蘑子实体的酪氨酸酶活性的第1个峰值均出现在贮藏6 d时。贮藏9~18 d,半球期和平展期采收的巨大口蘑子实体的酪氨酸酶活性均维持在较低水平,且与卷边期采收的巨大口蘑子实体的酪氨酸酶活性存在显著性差异(p<0.05)。贮藏18 d 后,原基期、卷边期、衰老期采收的巨大口蘑子实体的酪氨酸酶活性相对较高,且一直到贮藏30 d 结束时,这3 个发育期采收的巨大口蘑子实体的酪氨酸酶活性均显著高于其他5 个发育期采收的巨大口蘑子实体,而其他5 个发育期采收的巨大口蘑子实体的酪氨酸酶活性差异不显著(p>0.05)。经分析,以上结果与原基期、卷边期、衰老期采收的巨大口蘑子实体的褐变度高,而其他发育期采收的巨大口蘑子实体的褐变度较低这一结果相吻合。
2.4 不同发育期采收的巨大口蘑子实体的漆酶活性变化情况
由表4可知,除了原基期和菇蕾期采收的巨大口蘑子实体之外,其他发育期采收的巨大口蘑子实体的漆酶活性,均随贮藏时间的延长呈波浪式下降的趋势。贮藏3~6 d,原基期、幼菇期、菇蕾期采收的巨大口蘑子实体的漆酶活性出现第1 个峰值。贮藏0~18 d,卷边期、半球期、平展期采收的巨大口蘑子实体的漆酶活性迅速下降。贮藏15 d 时,幼菇期采收的巨大口蘑子实体的漆酶活性较高,菇蕾期采收的巨大口蘑子实体的漆酶活性较低,其他发育期采收的巨大口蘑子实体的漆酶活性均处于两者之间。贮藏18 d 后,卷边期、半球期采收的巨大口蘑子实体的漆酶活性开始缓慢上升,而平展期采收的巨大口蘑子实体的漆酶活性在贮藏21 d后开始缓慢上升。贮藏0~18 d,半球期采收的巨大口蘑子实体的漆酶活性波动较大,并在贮藏18 d后漆酶活性一直处于下降状态。中菇期采收的巨大口蘑子实体的漆酶活性在整个贮藏期内下降较为平缓,并在贮藏30 d 结束时下降到最低。经分析,以上结果与原基期、卷边期、衰老期采收的巨大口蘑子实体的褐变度高,而其他发育期采收的巨大口蘑子实体的褐变度较低这一结果相吻合。

表4 不同发育期采收的巨大口蘑子实体的漆酶活性变化情况
2.5 不同发育期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性变化情况
由表5可知,不同发育期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性,均随贮藏时间的延长呈波浪式变化的趋势,且波动相对恒定。不同发育期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性在贮藏3~9 d、12~18 d、21~27 d,均出现了不同的峰值。经分析可知,不同发育期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性的峰值均出现在总酚含量的峰值之前,这表明巨大口蘑子实体苯丙氨酸解氨酶活性的升高,启动了巨大口蘑子实体总酚的合成,苯丙氨酸解氨酶是酚类底物合成的关键酶。
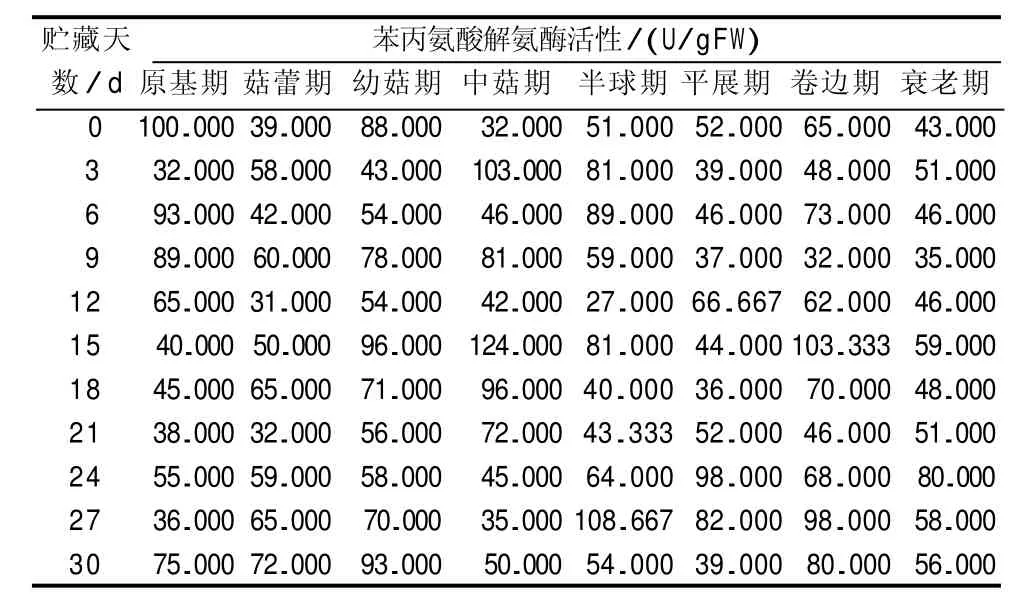
表5 不同发育期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性变化情况
2.6 不同发育期采收的巨大口蘑子实体的总酚含量变化情况
由表6可知,不同发育期采收的巨大口蘑子实体的总酚含量,均以刚采摘时的总酚含量为中心呈波浪式变化的趋势,且波动相对恒定。其中,卷边期采收的巨大口蘑子实体的总酚含量变化幅度最小,且含量一直维持在较低水平,与原基期、菇蕾期、幼菇期采收的巨大口蘑子实体的总酚含量存在显著性差异(p<0.05)。原基期、幼菇期、中菇期、平展期采收的巨大口蘑子实体的总酚含量在贮藏9 d 时出现了第1 个峰值,半球期采收的巨大口蘑子实体的总酚含量在贮藏12 d时出现第1个峰值,卷边期、菇蕾期采收的巨大口蘑子实体的总酚含量在贮藏15 d 时出现第1 个峰值。整个贮藏期间,原基期、菇蕾期、幼菇期、中菇期采收的巨大口蘑子实体的总酚含量相对较高,半球期、平展期、卷边期、衰老期采收的巨大口蘑子实体的总酚含量相对较低,这表明幼嫩子实体的代谢旺盛,酚类物质合成量相对较高。

表6 不同发育期采收的巨大口蘑子实体的总酚含量变化情况
2.7 不同发育期采收的巨大口蘑子实体的褐变度与各生理指标的相关性分析
相关性分析是在控制其他变量的线性影响下,分析两个变量间的线性关系。由表7可知,不同发育期采收的巨大口蘑子实体的褐变度与多酚氧化酶活性均呈负相关,特别是原基期、菇蕾期、幼菇期、平展期、卷边期、衰老期采收的巨大口蘑子实体的褐变度与多酚氧化酶活性呈极显著负相关(p<0.01);除了原基期、衰老期采收的巨大口蘑子实体之外,其他6 个发育期采收的巨大口蘑子实体的褐变度与酪氨酸酶活性呈负相关;除了原基期、菇蕾期采收的巨大口蘑子实体之外,其他6 个发育期采收的巨大口蘑子实体的褐变度与漆酶活性呈负相关;除了原基期、中菇期采收的巨大口蘑子实体之外,其他6个发育期采收的巨大口蘑子实体的褐变度与苯丙氨酸解氨酶活性呈正相关;除了原基期采收的巨大口蘑子实体之外,其他7 个发育期采收的巨大口蘑子实体的褐变度与总酚含量呈负相关。

表7 不同发育期采收的巨大口蘑子实体的褐变度与各生理指标的相关性分析
3 结论与讨论
试验结果表明,贮藏15 d 内,不同发育期采收的巨大口蘑子实体的褐变度变化相对较平缓,贮藏15 d 后,不同发育期采收的巨大口蘑子实体的褐变度开始缓慢上升,且子实体的褐变度与多酚氧化酶活性、酪氨酸酶活性的相关度高。例如,原基期、卷边期、衰老期采收的巨大口蘑子实体的褐变度高,主要是由于其多酚氧化酶、酪氨酸酶活性高,促进了子实体中酚类物质的氧化(原基期采收的巨大口蘑子实体的总酚含量和苯丙氨酸解氨酶活性相对较高也是其褐变度高的原因之一),而半球期、平展期采收的巨大口蘑子实体的褐变度低,主要是由于其多酚氧化酶、酪氨酸酶活性低,自身的氧化反应也较少发生,这说明多酚氧化酶、酪氨酸酶活性低是巨大口蘑子实体不易褐变、耐贮藏的重要原因。同时,在整个贮藏期间,半球期、平展期、卷边期、衰老期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性变化幅度较小,波动相对恒定,这就使酚类底物合成反应发生缓慢,总酚含量较少;而原基期、幼菇期、中菇期采收的巨大口蘑子实体的苯丙氨酸解氨酶活性变化幅度相对较大,这就使底物合成反应发生旺盛,总酚含量较高。
在本试验中,不同发育期采收的巨大口蘑子实体的多酚氧化酶活性,均随贮藏时间的延长大体上呈波浪式下降的趋势,该结果与杨春敏等[18]、冯立娟等[24-25]的研究结果一致,但与韩春然等[26]、张晓聪等[27]的研究结果不同。同时,巨大口蘑子实体的多酚氧化酶活性低于双孢蘑菇、平菇、香菇等[28],表明多酚氧化酶活性的差异是造成巨大口蘑子实体相比其他食用菌不易褐变的主要原因。除了原基期和菇蕾期采收的巨大口蘑子实体之外,其他发育期采收的巨大口蘑子实体的漆酶活性,均随贮藏时间的延长呈波浪式下降的趋势,该结果与喻晓明等[29]的研究结果一致。
综上所述,巨大口蘑子实体的适宜采收时期为半球期和平展期。
