草地贪夜蛾RACK1基因生物信息学及表达分析
2023-03-27严哲钰张尚志
何 凯,邓 凡,严哲钰,张尚志,2
(1.湖南人文科技学院农业与生物技术学院,湖南 娄底 417000;2.湖南省农田杂草防控技术与应用协同创新中心,湖南 娄底 417000)
草地贪夜蛾(Spodoptera frugiperda)是典型的鳞翅目害虫,可危害作物超过350 种,主要取食以禾本科为主的农作物,严重损害寄主作物的产量和品质[1]。草地贪夜蛾的适应能力比较强,同时迁移速度快,繁殖能力强。自2019 年草地贪夜蛾进入我国云南地区以来,已在我国多个省份出现,给农业生产带来较大损失,严重影响我国农业的可持续发展[2]。目前,关于草地贪夜蛾相关的研究主要集中在其发生发展及防治技术方面,但其分子生理尤其是与病毒侵染相关的分子机制研究相对较少。
蛋白激酶C 受体l(Receptor for Activated C Kinase1,RACKl)是细胞内一种高度保守的衔接蛋白,在原核生物和真核生物中都有广泛的表达[3]。RACK1蛋白于1991年被Daria首先用试验证明存在,直到1994 年才被Ron 等[4]克隆表达出来。研究表明,RACK1 是一种穿梭蛋白,其作为PCK 的受体,在生命体内发挥了重要作用[5]。目前已有研究发现,RACKl 参与到细胞分化、生长和凋亡以及胚胎发育等多个生命活动,并且均起到重要的调控作用。
目前已有多项研究证实RACK1 在病毒入侵过程中扮演着重要的角色。在MARC-145 细胞中,当干扰RACK1 表达后,猪生殖和呼吸道综合症病毒(Porcine reproductive and respiratory syndrome virus,PRRSV)的复制受到明显的抑制[6]。家蚕RACK1蛋白能与家蚕质型多角体病毒(Bombyx moricypovirus,BmCPV)上的蛋白结合,在病毒入侵过程中起着非常重要的作用[7],同时也有研究表明家蚕RACK1 蛋白能与家蚕核型多角体病毒(Bombyx morinucleopolyhedrovirus,BmNPV)互作,暗示其可能在BmNPV 入侵过程中扮演着非常重要的角色,但具体的机制还有待深入研究[8]。另外,研究显示,RACK1 能够与VDAC2 以及鸡传染性法氏囊病病毒(Infectious bursal disease virus,IBDV)VP5 蛋白相互作用,形成一个复合体,通过抑制细胞的凋亡从而促进病毒的复制,表明RACK1 在IBDV 侵染过程中起着关键作用[9]。
目前,RACK1 在多种生物体中的相关功能都已被研究报道,然而,迄今为止关于草地贪夜蛾RACK1 的相关研究暂无报道。课题组前期从转录组数据中筛选获得了草地贪夜蛾的RACK1 序列,该研究通过生物信息软件对草地贪夜蛾RACK1 进行生物信息学分析,同时分析RACK1 在草地贪夜蛾不同发育阶段不同组织以及NPV 刺激后不同时间段的表达模式,探究草地贪夜蛾RACK1 的功能以及该基因是否应答NPV 刺激,为今后RACK1 应答NPV 侵染的深入研究及其生物防治提供依据。
1 材料与方法
1.1 试验材料
草地贪夜蛾由西南林业大学徐进教授惠赠。苜蓿银纹夜蛾核型多角体病毒(AcMNPV)由实验室前期保存,繁殖。高保真酶、DNA 纯化回收试剂盒、质粒小提试剂盒,克隆菌株Escherichia coli DH5α及pMD19-T 载体购自北京擎科生物科技有限公司;cDNA 反转录试剂盒、TB Green Premix Ex Taq II 购自宝生物工程(大连)有限公司。
1.2 试验方法
1.2.1 草地贪夜蛾的饲养及材料收集草地贪夜蛾每头幼虫单独饲养,用新鲜玉米叶喂食,在卵、幼虫、蛹、蛾的时期收集材料。同时,在六龄期解剖幼虫,收集各个组织,用DEPC 水清洗,去除杂质和其他组织,置于离心管中,液氮冷冻,放入-80℃冰箱保存。
1.2.2 草地贪夜蛾接种病毒随机挑选六龄草地贪夜蛾60 头,再随机平分为2 个组,每个组(试验组和对照组)又分别设置3 个技术重复和生物学重复。将幼虫饥饿处理数小时后,试验组每头经口添食5 μL AcMNPV 病毒液(1×107个/mL),对照组经口添食5 μL ddH2O,处理后同时给予玉米叶。在添毒后的不同时间点(6、12、24、48 和72 h),解剖幼虫收集中肠组织,用DEPC 水清洗,去除杂质和其他组织,置于离心管中,液氮冷冻,放入-80℃冰箱保存。
1.2.3 RNA的提取、cDNA合成、引物设计及克隆将病毒侵染后不同时间段收集的中肠及其他组织样品根据试剂盒说明书提取RNA,并合成cDNA,具体步骤同前期方法[10]。根据实验室前期转录组数据,从中筛选出具有完整开放阅读框的RACK1 进行研究。利用PrimerPremier5.0 软件设计基因克隆引物SfRACK1-F:5' -ATGACTGAAACATTGAA GCTT-3';SfRACK1-R:5' -TTATCGTGCTGAGACTGACAC-3';引物均由北京擎科生物科技有限公司合成。PCR反应条件:95℃ 5 min;94℃30 s,52℃ 30 s,72℃45 s,35 个循环;72℃ 10 min。利用琼脂糖凝胶电泳对PCR 产物进行检测,目的片段通过胶回收试剂盒回收,回收产物连接至pMD19-T 载体并转化到克隆菌株Escherichia coli DH5α 感受态细胞中,挑选20 个经蓝白斑筛选后的阳性克隆,经菌液PCR 验证后,将菌液送上海生工生物工程有限公司进行测序。
1.2.4 RT-qPCR根据SfRACK1核苷酸序列设计出荧光定量引物:SfRACK1-F:5'-CTGACC AGGGACGA AACCAA-3';SfRACK1-R:5'-CTGAGAAAGCCACGGA GA-3'。GG 内参引物为SfGADPH-F:5'-CGTGGGAG CAGGTTTCGT-3';SfGADPH-R:5'-ACGGCCGCT TTAGTG TTGTC-3'。RT- qPCR 具体步骤参照TaKaRa生物公司荧光定量试剂盒说明书,在8 连管中加入总体积为25 μL 反应液,通过CFX96TM Real-Time PCR Detection System 仪器进行RT-qPCR,程序为:95℃预变性30 s;扩增条件为95℃ 5 s,60℃ 30 s,40 个循环;溶解曲线95℃ 1 min;55℃ 30 s。获得数据通过2-△△Ct 的方法进行分析[11],并通过SPSS软件对各组数据的差异显著性进行分析。
1.2.5 SfRACK1生物信息学分析通过在线网站SignalP5.0 Server(http://www.cbs.dtu.dk/services/SignalP)和SMART(http://smart.embl-heidelberg.de)对SfRACK1 蛋白的信号肽及保守结构域进行预测;再通过在线网站 ExPASy(http://web.expasy.org/com pute_pi/)对SfRACK1 蛋白相对分子质量及等电点进行预测;通过TMHMM2.0(http://www.cbs.dtu.dk/services/TMHMM/)和Prot Scale(https: //web.expasy.org/protscale/)分别预测蛋白的跨膜结构和疏水性;利用在线工具SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)和SWISS-MODEL(https://swissmodel.expasy.org/interactive)分别对其二级结构和三级结构进行预测。采用MEGE6 软件构建RACK1 蛋白的系统进化树。
2 结果与分析
2.1 SfRACK1的克隆及序列分析
通过PCR 扩增及其产物的测序成功克隆获得SfRACK1的ORF 序列(图1),测序结果显示其ORF 包含有960 bp 碱基,总共编码319 个氨基酸(图2 A)。通过ExPASy 在线网站预测其蛋白分子量为35.9 KDa,等电点为8.07。通过在线网站SMART对SfRACK1 蛋白保守结构域进行预测,结果显示SfRACK1 含有7 个重复的WD40 结构域(图2 B)。
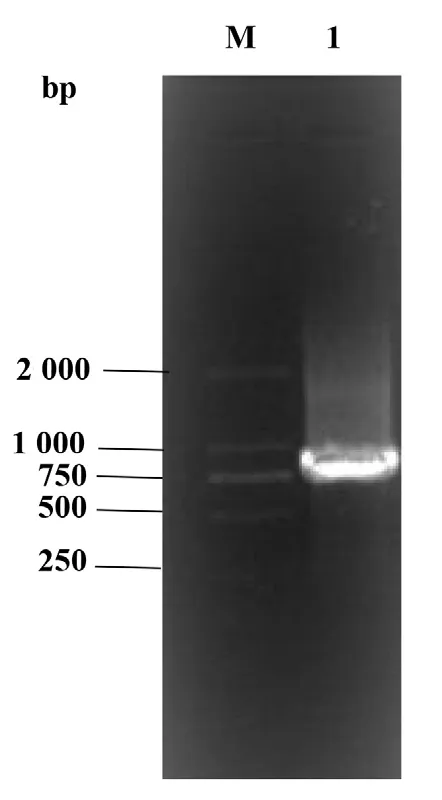
图1 SfRACK1基因PCR 扩增产物电泳图

图2 SfRACK1核苷酸和编码氨基酸序列分析
2.2 SfRACK1 信号肽、跨膜结构及亲疏水性分析
通过在线网站SignalP5.0 Server 对SfRACK1 蛋白的信号肽进行预测,结果显示SfRACK1 无信号肽(图3)。通过TMHMM2.0 在线预测蛋白是否存在跨膜结构,结果如图4 所示,SfRACK1 为膜外蛋白,无跨膜结构。利用 ProtScale 在线软件预测SfRACK1 蛋白的疏水性,结果表明,其氨基酸序列的预测分值最大为1.767,最小值为-2.722,平均值为-0.358,小于0,表明 SfRACK1 为亲水蛋白(图 5)。
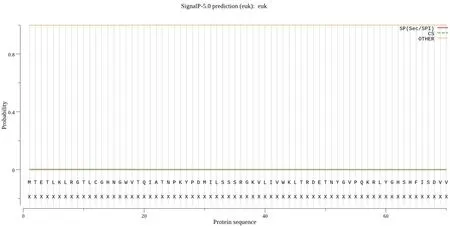
图3 SfRACK1 信号肽预测
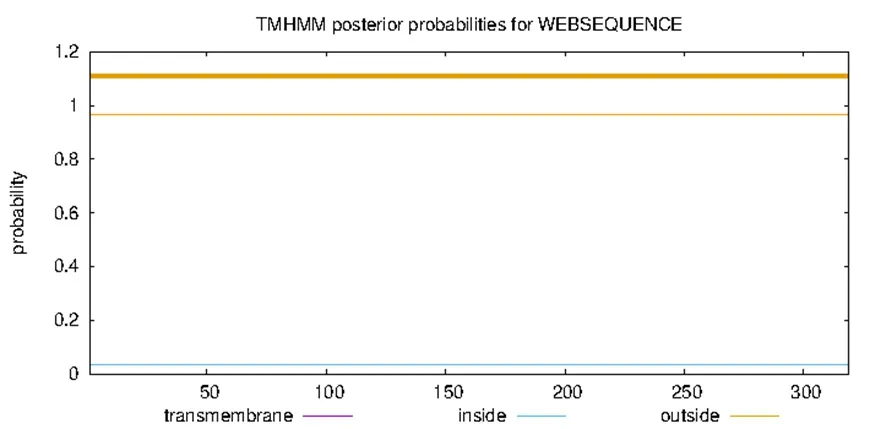
图4 SfRACK1 蛋白跨膜结构预测
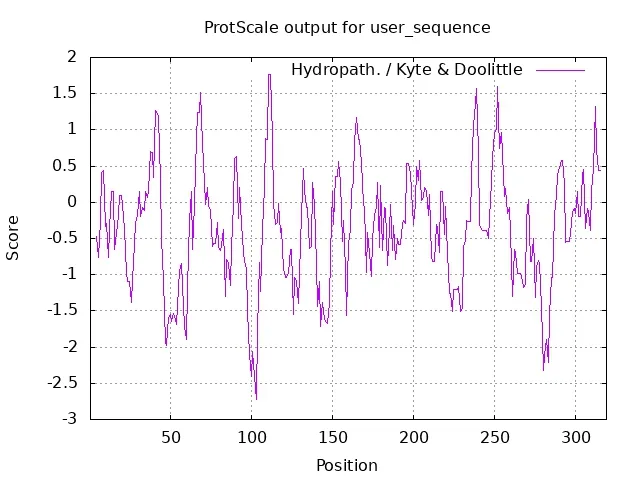
图5 SfRACK1 蛋白亲疏水性预测
2.3 SfRACK1 蛋白的二级结构及三级结构分析
通过在线工具SOPMA 对SfRACK1 蛋白的二级结构进行预测,结果(图6)表明,SfRACK1 蛋白二级结构主要由延伸链区(Extended strand 43.26%)、无规则卷曲(Random coil 37.30%)、β-转角(Beta turn 12.23%)和α-螺旋(Alpha helix 7.21.%)组成。SfRACK1 三级结构预测结果(图 7)显示,其为一个圆形结构,主要由无规则卷曲构成,这与前面二级结构所预测的结果一致。
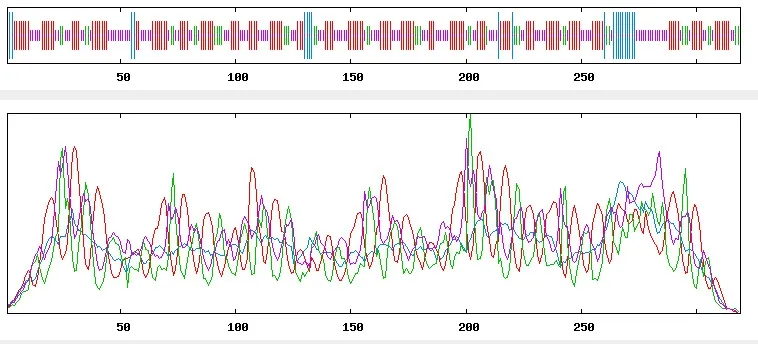
图6 SfRACK1 蛋白的二级结构
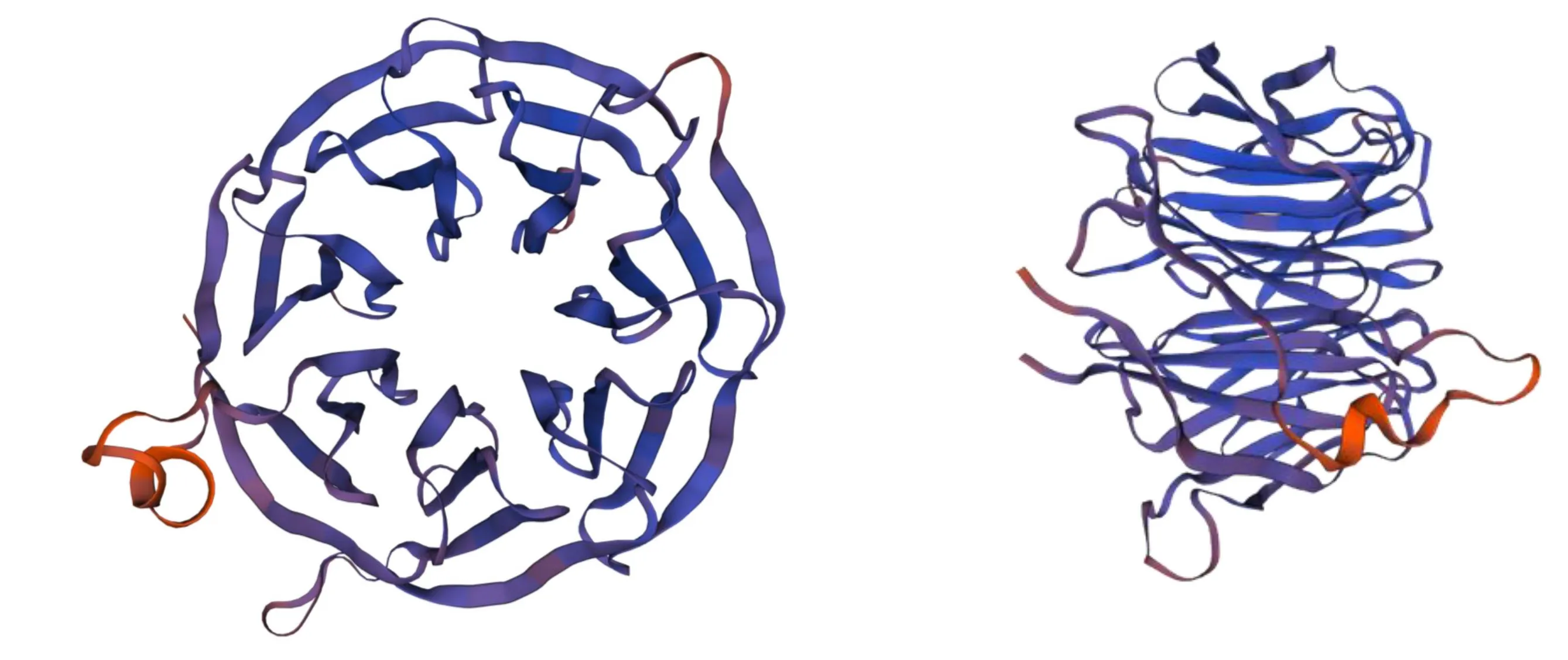
图7 SfRACK1 蛋白的三级结构
2.4 SfRACK1 与其他物种 RACK1 氨基酸序列比对及进化树构建
利用DNAMAN 软件对比SfRACK1 与其他物种氨基酸序列,由图8 可知,草地贪夜蛾与其他物种的RACK1 氨基酸序列具有较高的同源性,且与鳞翅目昆虫棉铃虫、小菜蛾等的同源性最高。为了调查SfRACK1 与其他昆虫的进化关系,该研究运用进化树来阐明它们的亲缘关系,结果(图9)显示,SfRACK1 与鳞翅目昆虫的RACK1 具有较高的同源性。

图8 SfRACK1 与部分物种 RACK1 氨基酸序列比对

图 9 以氨基酸序列构建的RACK1 系统进化树
2.5 SfRACK1基因的组织及时空表达分析
为分析SfRACK1在草地贪夜蛾不同组织及不同发育时期的表达水平,运用RT-qPCR 技术来检测SfRACK1的表达水平,结果如图10 所示,SfRACK1 在脂肪体中表达量最高,在头部、中肠中也具有相对较高的表达水平,但在血淋巴及表皮中表达量较低;在不同发育时期中,SfRACK1 在成虫期表达量最高,在卵期表达量最低。
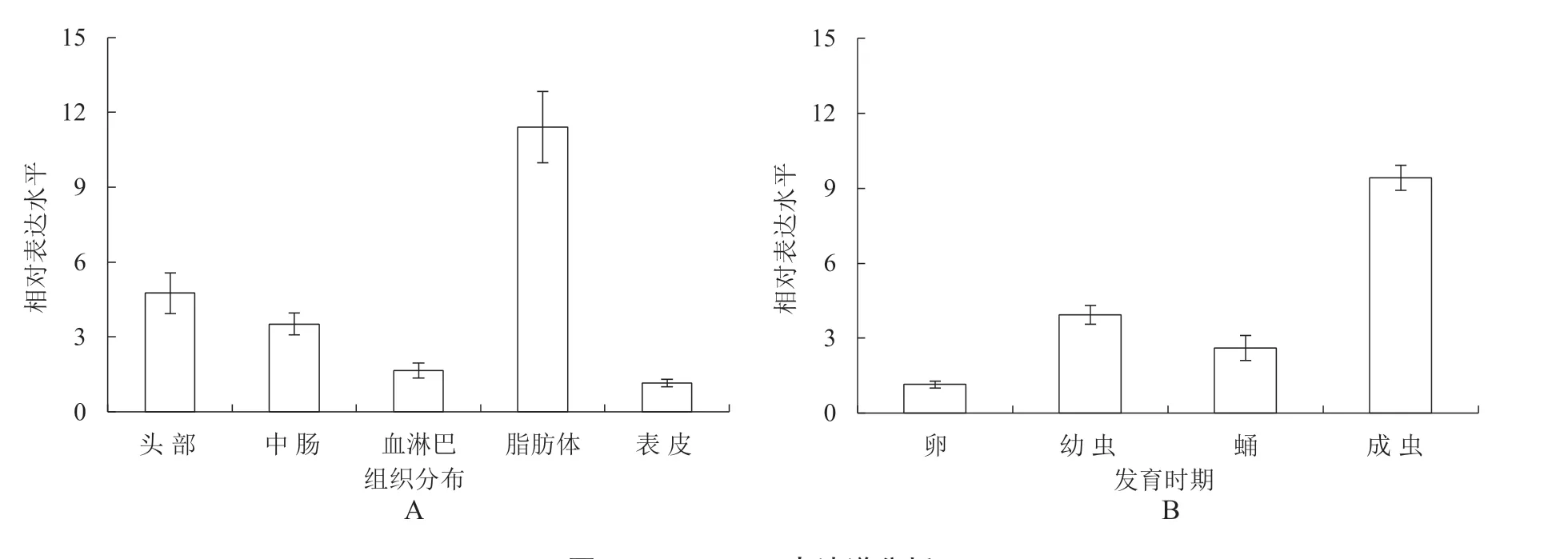
图10 SfRACK1表达谱分析
2.6 SfRACK1响应NPV 感染的表达模式
为探究SfRACK1是否能响应NPV 的侵染,运用RT-qPCR 方法从转录水平分析草地贪夜蛾受到NPV 侵染后不同时间点SfRACK1在中肠组织的表达模式,结果(图11)表明,与对照组相比,草地贪夜蛾受到NPV 侵染后不同时间点的SfRACK1表达量均有显著增加,且随着时间的延长而增加。这说明中肠中SfRACK1能够响应NPV 的侵染。
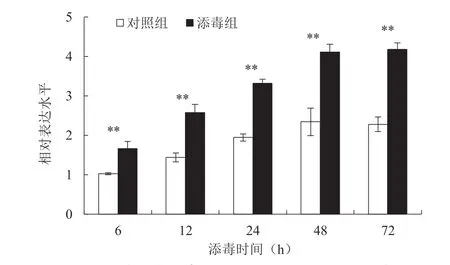
图11 NPV 侵染后草地贪夜蛾中肠SfRACK1的相对表达水平
3 结论与讨论
RACK1 是细胞内一种高度保守的衔接蛋白,在各种生物体中都有广泛的表达[3]。研究表明,RACK1 参与了生命体内细胞分化、生长和凋亡以及胚胎发育等多种生理活动[12]。笔者对草地贪夜蛾RACK1 的研究分析表明,SfRACK1基因的ORF 包含有750 bp 碱基,编码319 个氨基酸,相对分子质量为35.9 kDa,功能区包含有7 个WD40 结构域。多重序列比较发现SfRACK1 氨基酸序列与鳞翅目昆虫的氨基酸序列相似性均在90%以上。同时,通过构建系统进化树发现,SfRACK1 与鳞翅目昆虫RACK1 聚为一支。在家蚕[13]、柞蚕[14]等多种鳞翅目昆虫中RACK1 均已被鉴定,其分子质量均为36 kDa 左右,且具有高度的保守性。以上结果表明,SfRACK1 同其他鳞翅目昆虫RACK1 一样具有较高的保守性,暗示其可能具有相同的生物学功能。
RACK1 在草地贪夜蛾不同组织中的表达分析结果表明,SfRACK1 在脂肪体中具有较高的表达量,在中肠和头部表达量次之,而在表皮及血淋巴中表达量较低,这与其他昆虫RACK1 的表达模式基本一致。脂肪体作为昆虫先天免疫的主要场所,既是中间代谢组织,又是昆虫重要的免疫器官,脂肪体参与分泌合成了多种与分子免疫相关的蛋白[15]。而昆虫的中肠组织是昆虫抵御外来病原菌,尤其是食入性病原菌的第一道屏障[16-17]。已有研究证实,RACK1 能够参与应答病原菌的感染,SfRACK1 在脂肪体及中肠的高表达暗示着其可能参与了草地贪夜蛾的免疫作用。RACK1 在草地贪夜蛾不同发育阶段的表达分析结果表明,SfRACK1 在各个时期都有表达,但在成虫期表达量最高,在卵期表达量最低。这与小菜蛾[18]、家蚕以及柞蚕[14]中RACK1 的表达模式基本一致。RACK1 参与多个基本生物过程,涉及细胞生长和蛋白合成等[12]。在小菜蛾中通过RNAi 技术干扰RACK1 的表达可导致其死亡率上升、化蛹推迟、化蛹率降低以及蛹重减轻等问题。
草地贪夜蛾受到NPV 侵染后不同时间段中肠RACK1 的表达分析结果表明,与对照组相比,NPV侵染后其中肠的SfRACK1基因相对表达量显著提升。当柞蚕受到NPV 侵染后,RACK1 的表达水平也同样高于对照组[14];家蚕中不同抗性品系受NPV 侵染后,各品系的RACK1基因在中肠的表达水平也具有显著差异[8]。以上结果均表明,RACK1 能应答NPV的侵染,进而参与生物体的免疫反应。此外,已有研究表明,RACK1 蛋白能与BmCPV 上的蛋白结合,并且其表达水平的变化能影响病毒的入侵速度[7];同时,RACK1 蛋白还能与BmNPV 互作,但具体的作用及分子机制还有待深入研究[8]。
