贵州省鼠伤寒沙门菌单相变异株的CRISPR基因型和遗传多样性研究
2022-11-19白贵欢牟鸿江韦小瑜李世军刘淳婷汪俊华
白贵欢,游 旅,龙 利,牟鸿江,韦小瑜,,李世军,王 铭,刘 英,刘淳婷,王 丹,汪俊华
自20世纪90年代报道了一种与鼠伤寒沙门菌抗原式及遗传关系密切的沙门菌,即鼠伤寒沙门菌单相变异株,其是鼠伤寒沙门菌fljB基因突变或丢失从而导致II相鞭毛抗原(H2)无法表达的突变体[1],现已成为世界范围内一种非常重要的沙门菌血清型,在全球多个国家的畜牧、食品及人源样品中的检出率呈逐渐增高的趋势[2]。2020年郑州市从22家哨点医院收集分离的沙门菌中,鼠伤寒沙门菌单相变异株的检出率高于鼠伤寒沙门菌[3],在广东省珠海市该血清型成为本地婴幼儿腹泻病例中检出最多的菌株[4];2014—2018年江苏省食源性疾病监测中发现鼠伤寒沙门菌变异株的阳性率呈逐年增长的趋势[5]。近年,贵州省陆续从感染性腹泻病例中分离出鼠伤寒沙门菌单相变异株,而其基因型和遗传多样性尚不清楚。规律的成簇间隔短回文重复序列(Clustered Regularly Interspaced Short Palindromic Repeats,CRISPR)是由RNA介导的对外源核酸片段的干扰能形成获得性和可遗传性的生物免疫防御系统,广泛分布在细菌和古生菌的基因组中,由22~47 bp的重复序列和插入其中的间隔序列组成[6-7]。CRISPR的同向重复序列通常是保守的,间隔序列即使在同一种菌中,其重复数量和种类也可能各不相同,所以可作为靶标对细菌进行分型[8]。为了解贵州省鼠伤寒沙门菌单相变异株的基因型和遗传多样性特点,本研究采用CRISPR方法对分离自2013-2018年贵州省7个市(州)的71株鼠伤寒沙门菌单相变异株进行了CRISPR基因分型及遗传多样性分析,为贵州省鼠伤寒沙门菌单相变异株引起的沙门菌病的预防控制提供实验室依据。
1 材料与方法
1.1 菌株来源 基于贵州省感染性腹泻病例的常规监测,符合24 h腹泻3次及以上且伴有粪便异常(稀便、水样便、粘液便或脓血便)等的腹泻患者粪便标本进行鼠伤寒沙门菌单相变异株的分离培养鉴定。2013年1月至2018年12月共收集到71株疑似鼠伤寒沙门菌单相变异株,其中铜仁市30株、贵阳市9株、安顺市9株、遵义市17株、六盘水市1株、黔东南州3株、黔南州2株。
1.2 主要试剂及仪器 PCR扩增仪为德国80WTprofessional Gradient 96;Pre Mix和DNA Marker DL2000购自大连宝生物公司;凝胶成像仪购自美国Bio-Rad公司;引物由北京天一辉远生物公司合成。
1.3 菌株复核及血清分型 菌株接种至沙门菌显色培养基上,36℃培养过夜,然后挑取淡紫色菌落接种于克氏双糖铁培养基和动力-吲哚-尿素培养基上进行初步生化鉴定,再将符合沙门菌初步生化鉴定的菌株使用API20E生化鉴定板进行系统生化鉴定,对明确鉴定为沙门菌的菌株转种至营养琼脂平板36℃过夜培养进行O相抗原凝集,确定O相抗原后点种至沙门菌H相诱导培养基进行H1、H2相血清凝集,依据White-Kauffman抗原表结合O相、H1相和H2相血清凝集结果对沙门菌进行血清分型。
1.4 细菌DNA提取及双重PCR法鉴定菌株 将血清分型为鼠伤寒沙门菌单相变异株的菌株划线接种至血琼脂平板,36℃培养过夜,然后刮取适量菌苔制成菌悬液,金属浴100℃10 min,13000 r/min离心10 min,吸取上清液得到细菌DNA。以提取的DNA作为模板,引物参考Tennant等[9]的设计,PCR反应体系25μL,其中Pre Mix 12.5μL、上下游引物各1μL,模板DNA 1μL,用纯水补至25μL。PCR反应条件:95℃预变性2 min;95℃变性30 s,64℃退火30 s,72℃延伸90 s,共30个循环;最后72℃延伸10 min。PCR扩增产物在2%琼脂糖凝胶电泳后观察结果,出现1389 bp和1000 bp 2条扩增条带的判定为鼠伤寒沙门菌,仅出现1000 bp 1条扩增条带的判定为鼠伤寒沙门菌单相变异株。
1.5 CRISPR位点的检测及分析
1.5.1 引物合成 根据Fabre[10]及庄孝飞[11]的研究报道合成CRISPR1和CRISPR2引物,引物由北京天一辉远生物公司合成。
1.5.2 PCR扩增和测序 配制50μL PCR扩增体系:Pre Mix 25μL,上下游引物各2μL,DNA模板2 μL,纯水19μL,并分别对CRISPR1和CRISPR2位点进行PCR扩增。反应结束后,取5μL PCR扩增产物进行1%琼脂糖凝胶电泳,使用凝胶成像仪观察结果。将所得PCR扩增阳性产物送至北京天一辉远生物公司进行双向测序。
1.5.3 CRISPR序列的分析测序结果使用CRISPRCasfinder(https://crisprcas.i2bc.parissaclay.fr/Crispr CasFinder/Index)查找每个菌株的CRISPR位点,得到其重复序列及间隔序列。通过Weblogo(http://weblogo.berkeley.edu/logo.cgi)对重复序列进行保守性分析。根据Fabre[10]提出的命名方法对间隔序列进行命名和分型,将CRISPR1和CRISPR2位点的间隔序列分别命名为STM(Spacer of Typhimurium Monophase)和STMB,后缀B表示在CRISPR2位点中发现的间隔序列,间隔序列按发现顺序编号,对于每个分离株,CRISPR1和CRISPR22个位点的间隔序列组成一个CRISPR基因型,命名为TST(Typhimurium Sequence Type),不同型别用不同的数字后缀表示。进一步使用Bionumerics 8.0软件进行遗传多样性分析。
2 结果
2.1 菌株鉴定 71株疑似鼠伤寒沙门菌单相变异株经菌株复核、血清分型及双重PCR法鉴定均为鼠伤寒沙门菌单相变异株。
2.2 CRISPR位点检测 利 用PCR扩增菌 株的CRISPR1和CRISPR2位点,发现71株鼠伤寒沙门菌单相变异株中有56株菌检测到CRISPR1和CRISPR2位点,10株菌只检测到CRISPR1位点,5株菌只检测到CRISPR2位点。
2.3 重复序列组成及保守性分析 将测序结果与CRISPRCasfinder数据库进行比对,71株鼠伤寒沙门菌单相变异株共有2110个重复序列,其中CRISPR1位点有912个重复序列,CRISPR2位点有1198个重复序列。通过Weblogo软件分析结果如图1,菌株CRISPR位点的重复序列均包含高度保守 的、长29 bp的 共 有 序 列5′-CGGTTTATCCCCGCTGGCGCGGGGAACAC-3′,但CRISPR1位点的重复序列比CRISPR2位点的重复序列更为保守,具体表现为CRISPR1位点的重复序列共有17个碱基存在突变,CRISPR2位点的重复序列共有24个碱基存在突变。
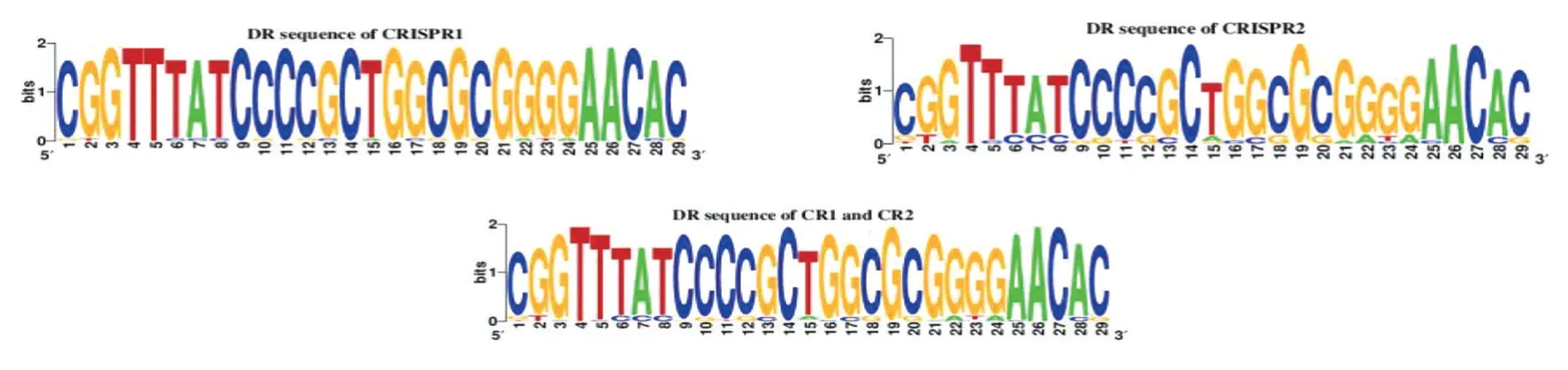
图1 71株鼠伤寒沙门菌单相变异株CRISPR位点重复序列的保守性分析Fig.1 Conservation analysis of CRISPR locus repeats of 71 monophasic Salmonella Typhimurium strains
2.4 间隔序列组成 71株鼠伤寒沙门菌单相变异株的CRISPR位点共检测到163个间隔序列(包含共享间隔序列),其 中CRISPR1有102个,CRISPR2有61个,长度为27~38 bp,未发现新的间隔序列。根据菌株CRISPR位点间隔序列组成,71株菌被分为19个CRISPR1型和11个CRISPR2型,基因型及出现频率如图2。CRISPR1位点的STM17间隔序列在部分菌株中重复2次,STM3、STM4、STM5、STM6、STM7、STM8这5个间隔序列存在于大部分菌株,在CRISPR2位点中,STMB8、STMB9、STMB10、STMB11、STMB12、STMB13、STMB14、STMB15这8个间隔序列存在于大部分菌株。

图2 CRISPR1和CRISPR2位点基因型的间隔序列组成及出现频率Fig.2 Spacer sequence composition and frequency of occurrence of CRISPR1 and CRISPR2 locus genotypes
2.5 CRISPR基因分型 将CRISPR1和CRISPR2位点的间隔序列联合分析(如图3),71株菌共得到33个CRISPR基因型(TSTs),命名为TST1-33,其中TST11和TST1是贵州省鼠伤寒沙门菌单相变异株的优势基因型,分别占总菌株数的25.35%和12.68%。除TST28和TST32、TST2、TST5、TST6、TST13、TST22、TST26、TST31只 检测出CRISPR1或CRISPR2位点从而组成独特的CRISPR基因型外,其余的TSTs间隔序列组成不同的CRISPR基因型。从CRISPR基因型的间隔序列组成来看,发现TST1和TST33虽间隔序列的排列相同,但由于TST33存在1个重复的间隔序列(STM17∗),从而将分离自贵阳市、遵义市、铜仁市和安顺市的菌株分成2个不同的CRISPR基因型。
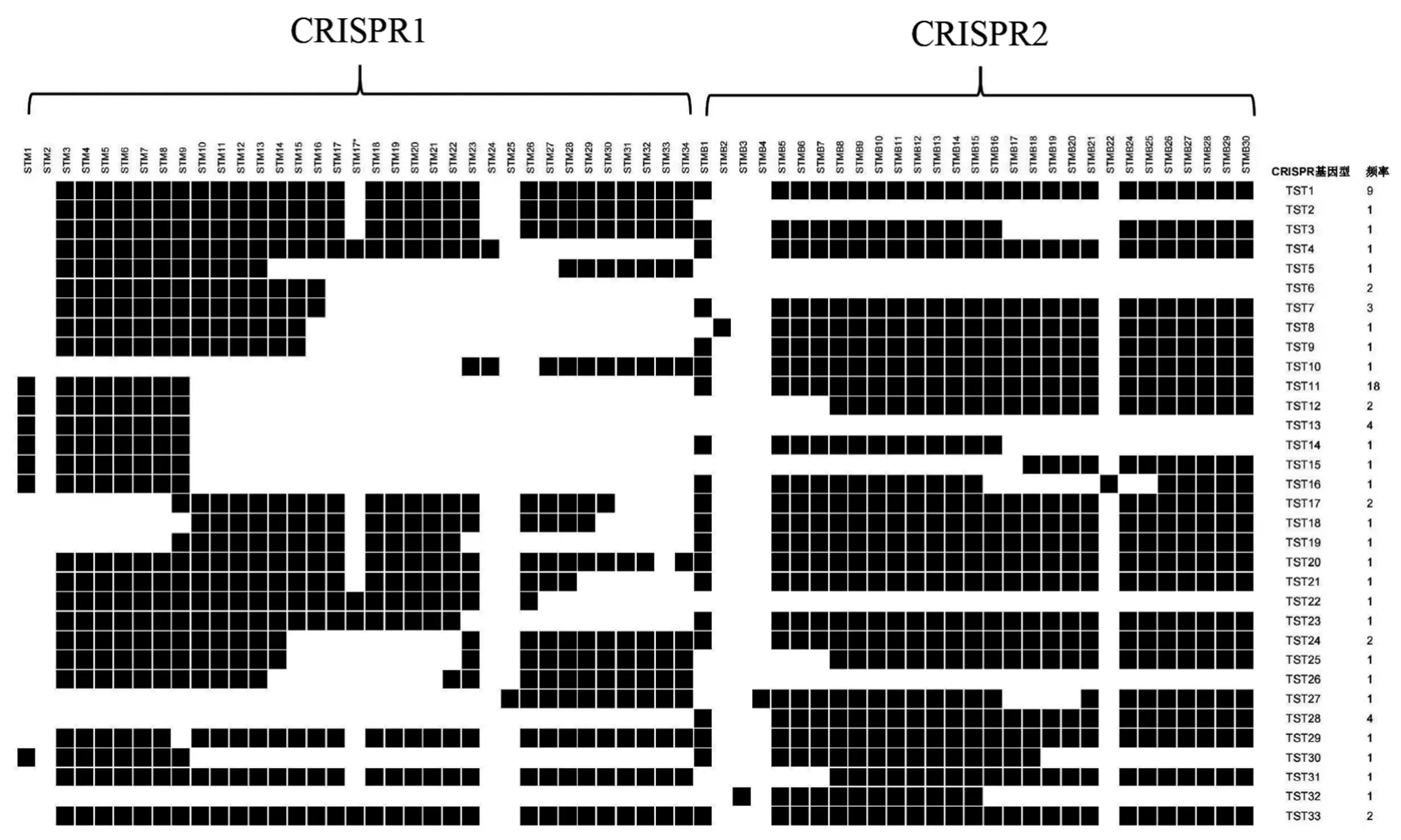
图3 贵州省71株鼠伤寒沙门菌单相变异株的33个TSTs基因型和出现频率Fig.333 TSTs genotypes and frequency of 71 monophasic Salmonella Typhimurium strains in Guizhou Province
2.6 聚类分析 使用BioNumerics软件对贵州省71株鼠伤寒沙门菌单相变异体与不同来源的参考株做系统进化树,结果见图4。根据各基因型亲缘关系的远近,33个TSTs基因型与3株不同来源的参考株被分为A、B、C、D 4个簇。A簇包含的菌株最多,主要来自2016—2018年的菌株,包括优势基因型TST11在内的12个TSTs基因型,来自贵州省7个市州的分离株。B簇包括另一个优势基因型TST1且包含的基因型最多(14个TSTs),分离自2013—2018年贵阳市、遵义市、安顺市、铜仁市和黔南州五个市州的菌株,其中TST33、TST1基因型的相似度达98.5%,2个基因型之间仅为1个间隔序列的差异。C簇由不同来源的3株参考株组成一个独立的分支。D簇包含7个TSTs型,来自铜仁市、遵义市、安顺市和黔东南州4个市州的分离株,但不含有2014年的菌株。整体来看,A簇CRISPR1、2位点的间隔序列排列差异不大,B、C两簇CRISPR1位点间隔序列较CRISPR2位点的间隔序列更为多样,而在D簇则大多缺失CRISPR2位点。

图4 贵州省71株鼠伤寒沙门菌单相变异株与3株不同来源参考株的系统进化树Fig.4 Phylogenetic tree of 71 monophasic Salmonella Typhimurium strains from Guizhou Province and three reference strains from different sources
3 讨论
沙门菌是常见人兽共患病病原菌,在世界各地的食物中毒病例中位居榜首,其引起的感染性腹泻给我国带来了严重的经济及社会负担[12]。鼠伤寒沙门菌单相变异株是鼠伤寒沙门菌的一种血清型,已被证明与鼠伤寒沙门菌具有相似的毒力和耐药特征[13],是目前常见的沙门菌血清型之一。在欧洲与人沙门菌病的常见血清型中,鼠伤寒沙门菌单相变异株在2017年排名第3,仅次于肠炎沙门菌和鼠伤寒沙门菌[14]。在我国河南省,鼠伤寒沙门菌单相变异株的检出率已超过鼠伤寒沙门菌,成为仅次于肠炎沙门菌排名第2的沙门菌血清型[15],在2018—2019年广东省中山市食源性疾病监测及食物中毒事件中,鼠伤寒沙门菌单相变异株是其优势血清型之一,目前已成为广东省最主要的流行血清型[16]。因此,及时、准确地对鼠伤寒沙门菌单相变异株进行鉴定、分型和进化分析等有重大意义。CRISPR作为一个细菌获得性免疫的机制,其重复序列具有高度的保守性,间隔序列通常来源于外源DNA,体现细菌在进化过程中的多样性,通过对不同菌株的间隔序列识别,可为细菌的鉴定、分型和进化分析等提供可靠的工作依据[17]。近年来对其研究逐渐深入。周松等[18]利用CRISPR分型技术将128株鼠疫菌分为2个基因型,发现Cb2型为河北省长爪沙鼠疫源地的主要基因型,谢晓蕾等[19]将不同来源的173株鼠伤寒沙门菌及单相变异株分为34个CRISPR基因型,其中TST4为优势基因型,经过溯源分析,发现猪是其优势基因型的主要宿主。另外,CRISPR分型还在空肠弯曲菌[20]、大肠杆菌[21]、志贺氏菌[22]等菌种有不同程度的应用。本研究使用CRISPR分型技术对分离自贵州省7个市(州)的71株鼠伤寒沙门菌单相变异株CRISPR位点重复序列的保守性、间隔序列组成、CRISPR基因型及遗传多样性进行了系统分析。
一般来说,沙门菌各血清型的重复序列显著保守,长度为29 bp,重复序列包含高度一致的序列5′-CGGTTTATCCCCGCTGGCGCGGGGAACAC-3′[10]。本 研 究 中 发 现,71株 菌CRISPR1和CRISPR2位点的重复序列与沙门菌CRISPR数据库中出现频率最高的重复序列一致,说明鼠伤寒沙门菌单相变异株CRISPR的重复序列具有高度的保守性。同时,在我们的研究中,CRISPR1和CRISPR2位点的重复序列分别有17和24个碱基存在突变,说明CRISPR2位点的重复序列碱基频率比CRISPR1位点的重复序列碱基频率要高,CRISPR1位点的重复序列比CRISPR2位点的重复序列更保守,与Shariat等[23]的研究结果一致,但在谢晓蕾[24]的研究中CRISPR1位点的重复序列仅存在4个碱基突变,CRISPR2位点的重复序列仅存在11个碱基突变,一方面可能是因不同血清型的沙门菌其重复序列碱基突变不同,另一方面也可能因菌株来源不同而导致。
有研究显示[25-26],CRISPR结构中的间隔序列被认为是截取和保留某些外源入侵基因的片段,因此其排列顺序反映了菌株对环境的适应性进化,而不同CRISPR的间隔序列是不同的,但相同的间隔序列则显示出其具有特异性及适应性,也有研究报道CRISPR间隔序列的多样性是由于单个间隔-重复单元或连续间隔-重复单元的复制或删除所形成的[10],因此该微变异导致了71株鼠伤寒沙门菌单相变异株被分为了19个CRISPR1型和11个CRISPR2型或33个CRISPR1-CRISPR2组 合型(CRISPR基因型),研究结果不仅说明CRISPR1位点间隔序列多样性较CRISPR2位点间隔序列多样性高,同时表明即使在相同血清型中,CRISPR基因型也具有高度的多样性,这对于细菌分型来说意义重大。本研究发现CRISPR1位点的STM17间隔序列在部分菌株中重复出现,可能原因是菌株被相似的噬菌体入侵或者是由于CRISPR序列的自我复制形成,其反映出它们在进化上可能有所关联,其机制有待我们进一步研究。同时发现TST1和TST33虽间隔序列排列相同,但由于TST33存在一个重复的STM17间隔序列,从而将来自不同地点的分离株分为不同的CRISPR基因型,说明CRISPR分子分型技术能在一定程度下区分同一血清型的变异。
PFGE分型技术被广泛的用于溯源,在日常监测或暴发事件中可以及时的发现污染源头,但该方法只能通过图谱分析条带信息,且实验操作繁琐,对实验人员要求较高,与PFGE相比,CRISPR分型方法不受实验室环境和操作人员的影响,通过观察间隔序列的排列顺序和碱基片段可直观的比较菌株的进化特点和碱基变异情况。本研究选取的3株参考株[10,19]分离自猪源、鸡源和人源的标本,包括来自湖南省、江苏省、江西省、辽宁省、浙江省、上海市和法国的分离株。从系统进化结果上看,33个TSTs型与3株参考株明显的分为独立的分支,在进化关系上遗传相似性不高,说明贵州省鼠伤寒沙门菌单相变异株与其他省份或国外的CRISPR基因型有较大差异,可能有其他的感染来源,推测可能受地域等因素影响,有待做进一步研究。从间隔序列的排列来看,不同来源的参考株有不同于贵州省33个TSTs基因型的间隔序列,这些间隔序列是否具有独特的地域性还需扩大菌株量进行深入研究。另外,本研究发现贵州省某些菌株与菌株间的遗传相似性较高,但菌株的分离地点多样。如TST12与优势基因型TST11的遗传相似性为94%,包含分离自铜仁市、贵阳市、遵义市、安顺市和黔南州5个市州的菌株,TST33、TST1、TST20和TST29这4个基因型的遗传相似性达98.1%,来源于贵阳市、遵义市、铜仁市和安顺市4个市州的分离株,表明贵州省鼠伤寒沙门菌单相变异株的CRISPR基因型具有遗传多样性,不同菌株CRISPR基因型之间的遗传关系密切,有可能来自相同的感染源。特别是作为优势基因型的TST11(存在于除2014年和2015年外的4年中)和TST1(在2013—2018年间均存在)同时包含贵阳市、铜仁市、遵义市和安顺市4个不同分离地点的菌株,提示我们TST11和TST1具有在不同分离地点相互传播的潜力且存在持续流行的情况,需引起重视。由于本研究收集的菌株量较少,不足以反映整体情况,将在以后的工作中结合流行病学资料进一步对该血清型进行系统监测,扩大菌株量对其进行深入研究。
利益冲突:无
引用本文格式:白贵欢,游旅,龙利,等.贵州省鼠伤寒沙门菌单相变异株的CRISPR基因型和遗传多样性研究[J].中国人兽共患病学报,2022,38(10):862-868,873.DOI:10.3969/j.issn.1002-2694.2022.00.136
