多糖和激素对珠芽魔芋多叶形成的调控作用
2022-09-14谭长龙李思雅琦年洪娟李昆志
谭长龙,李思雅琦,钱 澄,王 昊,年洪娟,李昆志
(昆明理工大学 生命科学与技术学院,云南 昆明 650500)
魔芋(Amorphophallus)是药食两用的草本类单子叶植物[1],已有2 000余年的种植历史,最早见于《本草纲目》《开宝本草》等古代典籍,由其球茎提取的葡甘聚糖在食品、医疗、化工等领域被广泛应用[2]。魔芋主产于中国的云南、贵州、四川、湖南、广西等省(自治区),日本、泰国、缅甸、印度、斯里兰卡等国家也均有分布[3-4]。魔芋现有记载品种200余种,在中国大范围推广种植的魔芋主要包括白魔芋(Amorphophallusalbus)、花魔芋(Amorphophalluskonjac)和珠芽魔芋(Amorphophallusbulbifer)3大类及其引种驯化品种[1]。其中珠芽魔芋泛指叶面生长气生珠芽的一类魔芋,其植株高大、抗病害能力强、繁殖系数高,其球茎一般2年生长即可成熟,是中国云南等高温多雨地区大力推广种植品种[5],极具科学研究和应用价值。与1年仅着生1片复叶的花魔芋相比,珠芽魔芋1年可连续生长4~7片叶,多叶生长导致其生长期延长,光合面积和光合时间增加,生长年限缩短[6-7]。
前人发现,魔芋多叶生长现象时有发生,例如张东华等[6]种植引自东南亚地区的珠芽魔芋品种发现,多数珠芽魔芋能自然生长2~3片叶,少数能萌生4片叶;李琳等[8]在云南临沧海拔1 200 m种植珠芽魔芋,发现种球能普遍长出5~7片叶。石新龙等[9]利用低温存储白魔芋球茎后出现多叶着生现象;刘海利等[10]使用不同浓度激素处理后能够促进花魔芋球茎生发多片侧叶;陆红叶[11]发现,不同浓度配比的外源激素能够调控珠芽魔芋叶片生长。前人通过研究模式植物拟南芥侧芽生长发现,调节植株葡萄糖含量变化可以控制生长素的运输而影响幼苗侧枝萌发生长[12];调控拟南芥细胞分裂素水平上升以及使用生长素抑制剂均可促进侧芽大量形成[13]。科研工作者对魔芋出现多叶生长现象一直颇感兴趣,但对于珠芽魔芋多叶形成机制目前尚未见详细报道。
本研究通过分析珠芽魔芋实生种子自然萌发幼苗各时期各器官还原性多糖、淀粉和吲哚乙酸(IAA)、玉米素(ZT)、赤霉素(GA3)、脱落酸(ABA) 4种植物内源激素含量的变化,以及外施ZT和IAA处理珠芽魔芋幼苗各时期激素含量的变化,以期阐明糖分和激素含量变化与珠芽魔芋多叶生长的关系,揭示珠芽魔芋多叶形成的机制,为魔芋的栽培提供理论依据。
1 材料与方法
1.1 栽培管理
试验材料为本实验室自繁珠芽魔芋(A.bulbifer)种子,对照(CK)为自繁花魔芋(A.konjac)种子,盆栽于昆明理工大学温室大棚(102°51′51″E,24°50′52″N,海拔1 907 m),盆栽土壤为第四纪红壤。
珠芽魔芋种子于2020年4月1日按照陆红叶[11]的方法浸种催芽,20 d后播种。每盆点播发芽种子6粒,共72盆,盆高30 cm,盆底直径35 cm,顶部直径50 cm,每5 d洒水1次湿润土壤。出苗后每3 d浇水1次,除播种前施入适量底肥外,整个生长期均不追肥,并保持盆栽表面无杂草。珠芽魔芋幼苗6月中旬开始出土生长第1叶,随后长出第2叶,于7月中旬第1叶完全展开,之后平均每30 d长成1片新叶。2020年盆栽试验中幼苗最多长出4片叶,至同年11月底幼苗老叶开始枯黄,至次年1月中旬幼苗植株开始倒伏。对照为栽种的24盆花魔芋,按照珠芽魔芋幼苗同等条件栽培管理。花魔芋幼苗5月中旬出土生长,至同年12月初开始枯萎倒伏,生长期间未见多叶分化。分别于7月15日、8月15日、9月15日、10月15日采集珠芽魔芋1叶期、2叶期、3叶期、4叶期植株和对照同时期植株样品,用纯水快速洗净,吸水纸擦干表面后取叶片、叶柄、球茎和根部样,并迅速置入液氮中,在30 min内转移至-80 ℃冰箱储藏,用于测定不同器官内源激素含量。同时统计出叶数、叶柄长(株高)、球茎鲜质量和根长。每处理重复取样3次。
2021年4月1日将同品系珠芽魔芋种子按照2020年的试验步骤催芽播种。每盆播种2行,每行3粒,共72盆,采用与2020年试验相同的措施栽培管理。根据2020年激素检测结果,确定外源喷施激素分别为20 μg/L ZT溶液和40 μg/L IAA溶液。2021年6月20日开始对魔芋喷施外源激素,上午08:00浇水1次,并在浇水1 h后对其中24盆幼苗喷施清水(H2O)作为对照(CK),剩余48盆幼苗分两列分别喷施20 μg/L ZT和40 μg/L IAA,每隔10 d再喷施1次,连续处理3次。喷施激素后,水分管理为每3 d浇水1次,直至珠芽魔芋幼苗枯萎倒伏。分别于1叶期、2叶期、3叶期、4叶期,观察不同激素处理和CK珠芽魔芋新叶出土时间。1叶期至4叶期,当各处理和CK幼苗新叶展开后,及时取嫩叶部分并储藏于-80 ℃冰箱,用于测定内源激素含量。
1.2 魔芋幼苗糖分含量的测定
1.3 魔芋幼苗内源激素含量的测定
1.4 数据处理与分析
采用Microsoft Office Excel 2007整理试验数据;采用SPSS Statistics 17.0进行单因素方差分析,显著水平α=0.05;采用GraphPad Prism 8软件绘图。
2 结果与分析
2.1 自然生长珠芽魔芋幼苗生长情况
2020年自然生长过程中,珠芽魔芋各生长时期叶片生长情况如图1所示,叶柄长、球茎鲜质量、根长的变化如图2所示。由图2可以看出,1叶期,自然生长过程中珠芽魔芋第1片叶叶柄长与对照差异不大,均较低;2叶期,第2片叶叶柄长高于第1片叶,但二者差异不显著;3叶期,第2片叶叶柄长显著高于第1片叶;4叶期,第3片叶叶柄长最大,其次为第2片叶,均显著高于第1片叶。对照(花魔芋)整个生长期仅着生1片叶,随着时间的增加,叶柄长显著增加。随着珠芽魔芋幼苗长出第2、3和4片叶,叶面积不断增加,光合作用增强,4叶期魔芋球茎鲜质量和根长较前3个时期均显著增加。

图1 珠芽魔芋各时期幼苗叶片生长情况Fig.1 Leaves of Amorphophallus bulbifer seedlings at different stages
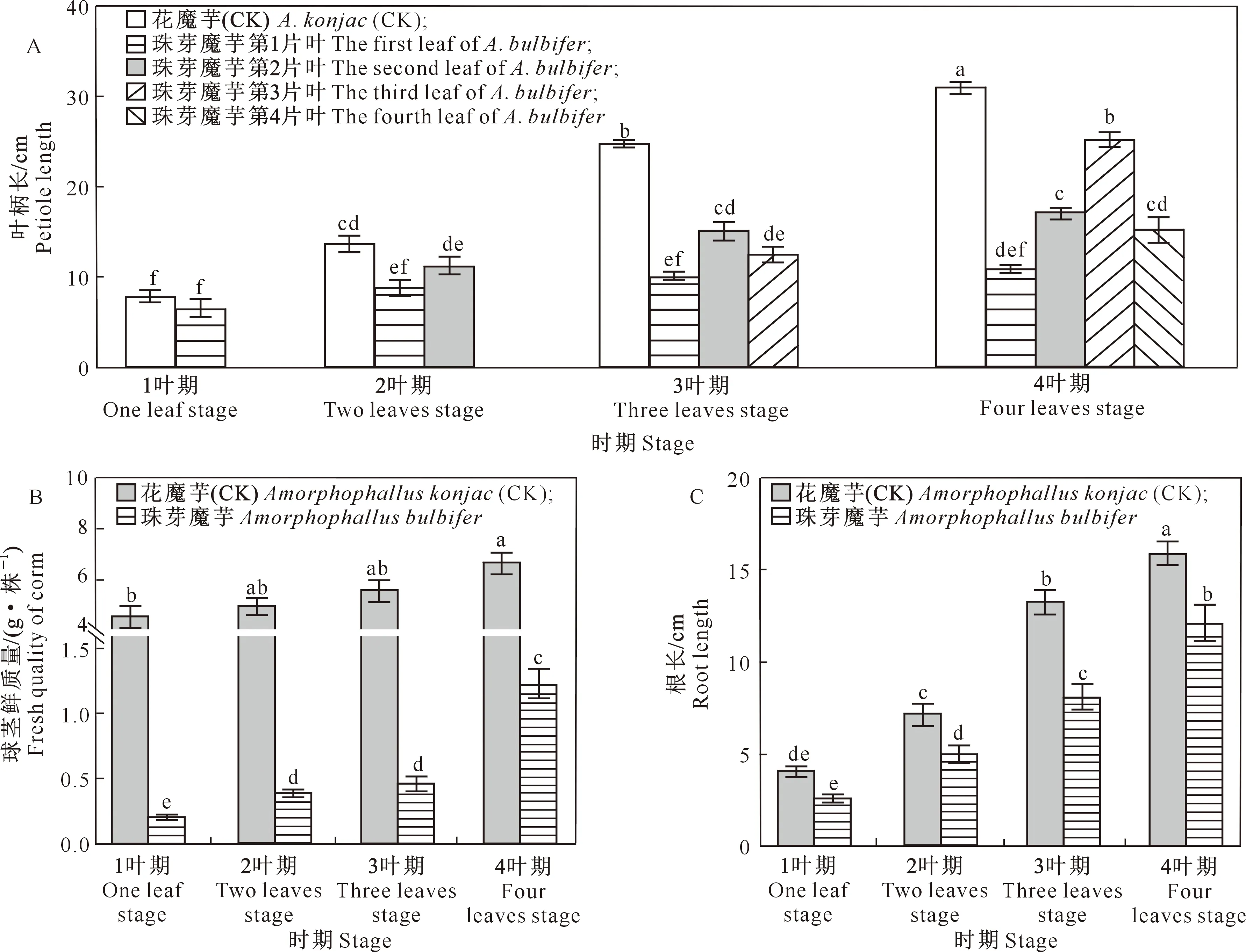
图柱上标不同小写字母表示差异显著(P<0.05)。图3~8同 Different lowercase letters indicate significant differences (P<0.05).The same forFig.3-8图2 各时期珠芽魔芋幼苗自然生长情况Fig.2 Natural growth of Amorphophallus bulbifer seedlings in different stages
2.2 自然生长珠芽魔芋幼苗糖分含量的变化
2020年,各时期自然生长珠芽魔芋幼苗不同器官还原性多糖和淀粉含量测定结果见图3和图4。由图3可以看出,与花魔芋相比,4个时期珠芽魔芋的叶片、叶柄、球茎和根中的还原性多糖含量均较高。由于光合作用旺盛且代谢缓慢,各时期珠芽魔芋第1叶停滞生长后,其叶片和叶柄中还原性多糖含量比同时期其他叶片高。各时期珠芽魔芋球茎还原性多糖含量较根中偏高,且两者还原性多糖含量随时间推移而增加。相较于老叶,各时期珠芽魔芋新叶(各时期最嫩叶片)还原性多糖含量均明显降低。这说明叶片光合作用合成的糖分通过叶柄运输至球茎并积累,促进球茎生长发育,老叶生长量低,还原性多糖含量较高,新叶生长量高,还原性多糖含量偏低。

横坐标中的‘1’表示1叶期,‘2’表示2叶期,‘3’表示3叶期,‘4’表示4叶期 。图4~8同 ‘1’ represents 1 leaf stage,‘2’ represents 2 leaves stage,‘3’ represents 3 leaves stage and ‘4’ represents 4 leaves stage.The same forFig.4-8图3 各时期珠芽魔芋幼苗自然生长状态下叶片、叶柄、球茎和根中还原性多糖含量的变化Fig.3 Changes of reducing polysaccharide content in leaf,petiole,corm and root of Amorphophallus bulbifer seedlings at different stages under natural growth state
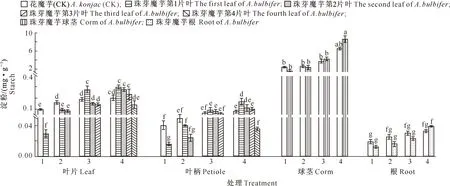
图4 各时期珠芽魔芋幼苗自然生长状态下叶片、叶柄、球茎和根中淀粉含量的变化Fig.4 Changes of starch content in leaf,petiole,corm and root of Amorphophallus bulbifer seedlings at different stages under natural growth state
由图4可以看出,在同一时期,自然生长的珠芽魔芋幼苗球茎中淀粉含量最高,其次是叶片和叶柄,根的最低。叶片、叶柄和根部淀粉含量均偏低,说明植株整体生长代谢旺盛,光合作用合成养分主要用于植株生长而不是积累储存。随着珠芽魔芋幼苗侧叶形成,整株光合作用合成糖分速率提高,导致各时期第1叶合成糖分开始累积并逐渐转化为淀粉,各时期第1片叶叶片和叶柄淀粉含量相较同时期其他叶片、叶柄高。在同一时期,相较于老叶,新叶淀粉含量均明显降低。这是因为侧芽萌发生长需要消耗大量糖分,当第2、3片叶长大并进行光合作用后,第1、2片叶中糖分运输减少,还原性多糖积累并开始转化为淀粉存储,相较新叶,老叶淀粉含量均明显上升。
2.3 自然生长珠芽魔芋幼苗激素含量的变化
采用高效液相色谱法检测各时期珠芽魔芋幼苗不同器官4种激素含量变化如图5~8所示。由图5可以看出,2020年,各时期自然生长珠芽魔芋幼苗叶片和根的ZT含量较高,而叶柄和球茎ZT含量较低,在同一时期叶片中,第1片叶ZT含量最高,且显著高于其他叶片。可知相较于老叶,各时期新叶ZT含量均明显降低。随着叶片数的增加,各时期叶柄和球茎的ZT含量总体降低,而根的ZT含量明显增加。随着生长期的延长,对照花魔芋叶片ZT含量明显增加,而叶柄、球茎、根的ZT含量无显著变化。

图6 各时期珠芽魔芋幼苗自然生长状态下叶片、叶柄、球茎和根中GA3含量的变化Fig.6 Changes of GA3 content in leaf,petiole,corm and root of Amorphophallus bulbifer seedlings at different stages under natural growth state
由图6可以看出,自然生长状态下珠芽魔芋幼苗各时期GA3含量以叶片中最高,其次是根,球茎中GA3含量最低。在各时期叶片中,第1片叶的GA3含量最低,其后新长出叶片的GA3含量均较高;叶柄的GA3含量变化与叶片的一致,即各时期新叶叶柄GA3含量较老叶均有所升高;各时期球茎和根的GA3含量之间无显著差异。花魔芋幼苗各时期不同器官GA3含量均显著低于珠芽魔芋,且各时期之间无显著差异。
由图7可以看出,各时期自然生长状态下珠芽魔芋幼苗IAA含量以根中最高,均明显高于叶片、叶柄和球茎。在叶片中,珠芽魔芋3叶期和4叶期的第3叶IAA含量显著高于同一时期的其他叶片;在叶柄和球茎中,各时期IAA含量差异变化不显著;4叶期根中IAA含量显著高于1叶期、2叶期和3叶期,而1叶期至3叶期之间IAA含量无显著差异。各时期叶片IAA含量差异显著,各时期花魔芋幼苗不同器官IAA含量均明显低于珠芽魔芋。
由图8可以看出,珠芽魔芋幼苗叶片、叶柄、球茎和根中ABA含量前期较低,后期较高,其中以4叶期的第4叶片、叶柄及其球茎和根的ABA含量最高。随着生长期的延长,花魔芋各器官的ABA含量无显著变化,且总体上低于珠芽魔芋。
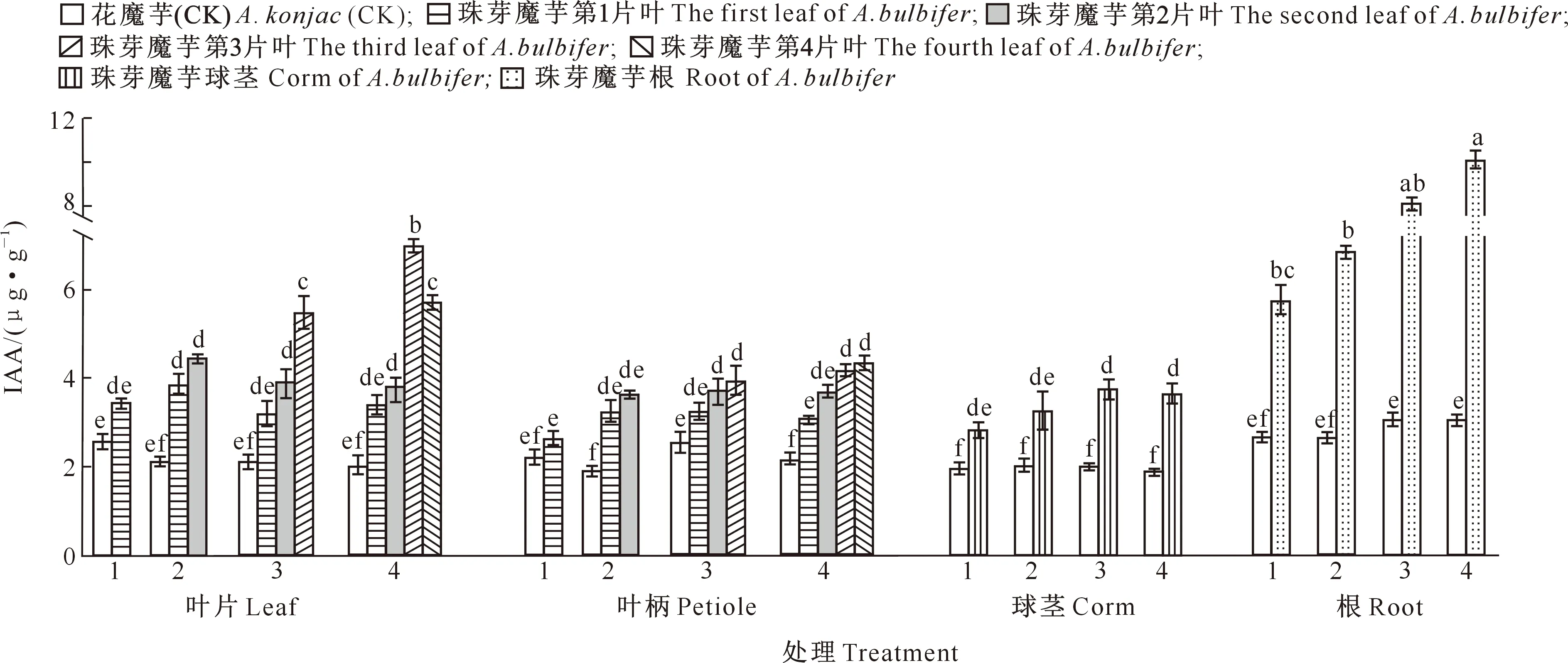
图7 各时期珠芽魔芋幼苗自然生长状态下叶片、叶柄、球茎和根中IAA含量的变化Fig.7 Changes of IAA content in leaf,petiole,corm and root of Amorphophallus bulbifer seedlings at different stages under natural growth state

图8 各时期珠芽魔芋幼苗自然生长状态下叶片、叶柄、球茎和根中ABA含量的变化Fig.8 Changes of ABA content in leaf,petiole,corm and root of Amorphophallus bulbifer seedlings at different stages under natural growth state
2.4 外施激素处理珠芽魔芋幼苗嫩叶激素含量的变化
依据2020年研究结果,可知各时期珠芽魔芋不同器官ZT和IAA含量变化差异较大,为此选择ZT和IAA作为外施激素处理珠芽魔芋幼苗。由表1可以看出,2021年, ZT处理和CK分别于8月28日和9月16日开始长出第4片叶,之后再无新的侧叶萌发;IAA处理最多只着生3片叶,相较CK和ZT处理出叶数减少。与CK比较,外施ZT能促进珠芽魔芋幼苗侧芽萌发速度;外施IAA推迟幼苗侧叶形成,抑制幼苗侧叶萌发。

表1 2021年不同激素处理珠芽魔芋新叶出土时间Table 1 Unearthed time of new leaf of Amorphophallus bulbifer with different hormone treatments in 2021
由图9可以看出,ZT处理幼苗各时期的ZT含量均明显高于CK和IAA处理,IAA处理幼苗各时期ZT含量均低于CK。相较CK,IAA和ZT处理的珠芽魔芋幼苗GA3含量均较高,且IAA处理的幼苗GA3含量高于ZT处理。IAA处理的珠芽魔芋幼苗各时期IAA含量均明显高于CK和ZT处理,ZT处理的的珠芽魔芋幼苗IAA含量与CK变化趋于一致。IAA处理的的珠芽魔芋幼苗各时期ABA含量均高于CK和ZT处理。
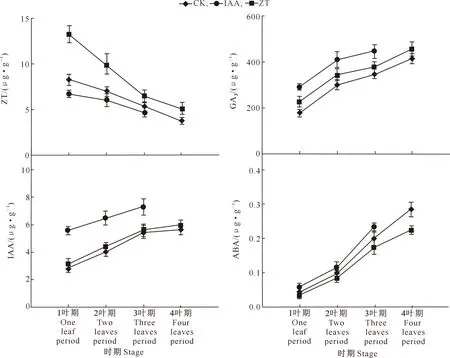
图例中‘IAA’和‘ZT’表示外施激素处理 ‘IAA’ and ‘ZT’ represent external hormone treatments图9 外源激素处理珠芽魔芋幼苗不同时期嫩叶激素含量的变化Fig.9 Changes of hormone content in young leaf of Amorphophallus bulbifer seedlings treated with exogenous hormones at different stages
3 讨 论
3.1 糖分对珠芽魔芋多叶形成的影响
植物的生长过程离不开糖分的调节,作为魔芋生长代谢的能量源泉,还原性多糖和淀粉在珠芽魔芋多叶形成过程中有着不可替代的作用。糖分是调控植株侧枝生长的一种重要信号物质[18],也是促进侧芽形成的重要因子,植株生长出现顶端优势一个重要因素就包括顶芽生长需要大量糖分并限制糖分转运到侧芽[19]。Stokes等[12]通过调节糖浓度诱导IAA运输及信号转导来调控拟南芥侧枝的萌发与生长。本研究发现,珠芽魔芋幼苗各个时期第2叶相较第3叶更容易萌发出芽,第3片叶相较第4叶更易出芽生长;随着生长期的延长,珠芽魔芋叶片和叶柄中还原性多糖含量逐渐升高,但各时期新叶叶片和叶柄中的还原性多糖含量低于老叶,说明较高的还原性多糖含量有利于植株生长壮大并促进侧叶萌发与生长。
作为植物光合作用累积合成的最终产物,淀粉含量变化是植株生长过程中一个重要指标。植株侧芽生长,需要消耗大量淀粉等物质[20]。本研究中,珠芽魔芋幼苗生长主要靠叶片光合作用提供养分,幼苗生长前期叶面积小,光合作用弱,合成有机物少,不足以转化成淀粉储存;随着珠芽魔芋侧叶生长,叶面积增大,光合作用增强,合成的有机物除了满足幼苗生长需求之外,盈余的有机物转化成淀粉储存于叶片和球茎中,且各时期老叶叶片和叶柄中的淀粉含量明显高于新叶。随着珠芽魔芋4叶期幼苗停止萌发生长新的侧芽,叶片光合产物大量累积并转化为淀粉,4叶期幼苗球茎中淀粉含量显著高于前3个时期,说明珠芽魔芋幼苗停止萌发侧芽前,植株不会大量合成淀粉储存。
3.2 激素对珠芽魔芋多叶形成的影响
内源激素是自然条件下植物生长发育的重要调节物质,其种类和数量的变化在珠芽魔芋多叶形成过程中起着重要作用。前人研究表明,侧芽ZT含量受植株顶芽控制[21],去除顶芽或施用生长素抑制剂能够上调异戊烯基转移酶(IPT)基因的表达,提高细胞分裂素含量,促进植物侧芽生长[22]。外施ZT可以增加植株侧芽数并促进侧枝生长[23]。本研究中,自然生长状态下珠芽魔芋幼苗各时期叶片和叶柄中ZT含量均以第1片叶和叶柄最高。各时期新叶ZT含量均显著低于老叶。与CK相比,外施ZT后,珠芽魔芋幼苗中ZT含量升高,侧芽萌发数量没有增加,但侧芽形成速度明显提高。可知外施适量ZT能促进珠芽魔芋ZT含量上升并促进侧叶的萌发和生长,说明珠芽魔芋中较高含量的ZT有利于侧叶萌发生长。
GA3作为一种常见的植物内源激素,在植株的生长过程中可以促进侧枝生长[24],并间接起到促进顶端优势的作用[25-26]。本研究中,随着生长期的延长,珠芽魔芋各时期叶片、叶柄、球茎和根部中GA3含量均逐渐上升,同时侧芽形成速度降低并逐渐停止萌发侧芽。
IAA作为最早被报道能维持植株顶端优势的激素,对植物侧枝萌发生长有着重要的调节作用[27]。前人研究发现,IAA的主要合成部位为顶芽和幼嫩的叶片[28],在植株顶芽去除部位外施IAA可以抑制侧芽生长[29]。对植株发芽原基施加生长素转运抑制剂N-1-萘基邻苯二甲酸 (NPA)和 2,3,5-三碘二苯甲酸 (TIBA) ,能引起生长素氧化基因双加氧酶(DAO)的上调,导致生长素水平迅速降低而诱导侧芽萌发[30]。本研究中,IAA处理珠芽魔芋IAA和GA3含量较CK和ZT处理均明显升高,但幼苗侧叶萌发形成速率降低,说明较高含量IAA和GA3不利于珠芽魔芋萌发侧芽。在自然生长状态下,珠芽魔芋幼苗期的叶片、叶柄、球茎中IAA含量长期处于较低水平,有利于侧叶形成,这是珠芽魔芋出现多叶生长现象的一个重要的因素。
ABA作为植物成熟和凋亡的重要调节激素,在植株侧枝形成过程中起着重要的调控作用。植株中ABA含量随侧芽的生长过程而发生变化[20],喷施ABA可以抑制侧芽生长[31]。在本研究中,随着生长期的延长,自然生长状态下珠芽魔芋各时期叶片、叶柄、球茎和根中ABA含量均逐渐上升,且各时期新叶ABA含量总体高于老叶。说明幼苗生长前期低含量ABA有利于珠芽魔芋萌发形成侧芽,较高含量的ABA抑制侧芽萌发并促进植株成熟。
4 结 论
随着生长期的延长,珠芽魔芋叶柄长、球茎鲜质量、根长总体呈增加趋势。珠芽魔芋糖分和内源激素含量随幼苗侧叶形成过程及其侧叶数量不同而发生了动态变化。在侧叶数量较少的幼苗初期,老叶中还原性多糖、淀粉和ZT含量较新叶均偏高。珠芽魔芋幼苗各器官中GA3、IAA和ABA含量均随着生长期的延长而逐渐升高。外施ZT能明显提高珠芽魔芋幼苗侧叶萌发速度,外施IAA明显抑制了珠芽魔芋幼苗侧叶萌发。较高含量糖分和ZT有利于珠芽魔芋侧叶萌发生长,较高含量GA3、IAA和ABA不利于珠芽魔芋侧叶萌发生长。
