冬季赤水河流域刚毛藻多样性调查及系统发育分析
2022-09-13赵志娟杨小霞钟俏君陈冠海彭相英刘本文刘国祥
赵志娟 杨小霞 钟俏君 陈冠海 彭相英 刘本文 朱 欢 刘国祥
(1. 南宁师范大学,北部湾环境演变与资源利用教育部重点实验室,南宁 530001;2. 南宁师范大学,广西地表过程与智能模拟重点实验室,南宁 530001;3. 中国科学院水生生物研究所,中国科学院藻类生物学重点实验室,武汉 430072)
刚毛藻(Cladophora)是水体中一类常见的丝状分枝绿藻,隶属于绿藻门石莼纲刚毛藻目(Cladophorales),在全球海洋及淡水中广泛分布[1,2]。其藻丝常着生于水中基质上或松散漂浮于水体表面,是水体中重要的生态类群,可为附植生物和底栖生物提供生产力和微生存空间[3,4]。
刚毛藻形态多为具分枝的丝状体,细胞管状多核,具网状周生色素体;藻体通过顶端或居间细胞分裂生长,通过基部细胞、假根或固着器进行固着。由于生态变异幅度较大、缺乏有效的形态诊断特征,该属的分类鉴定一直较为困难[5,6]。基于传统形态学方法对刚毛藻进行分类鉴定的结果正不断受到挑战[7,8]。
随着分子生物学技术的发展,基于多分子标记的序列分析越来越多地被用于刚毛藻目的系统分类中[9—11]。Boedeker等[10]在对全球刚毛藻科样品进行广泛调查和分析后提出: 在当前刚毛藻科物种形态分类可靠性较低、部分模式标本难以追溯确认及获得有效序列的情况下,依靠分子数据来推动该科的系统分类工作将是有帮助的。核糖体小亚基(SSU rDNA)和核糖体大亚基(LSU rDNA)是刚毛藻目系统分类研究中应用最为广泛的两个分子标记[12,13]。此外,内转录间隔区序列(ITS,ITS1-5.8S-ITS2)的使用也越来越多地受到重视。ITS序列相较于保守的SSU rDNA和LSU rDNA进化速率更快,因此更适用于解决近缘物种的系统分类[14—16]。Boedeker等[10]和Zhu等[11]关于刚毛藻目的研究均表明淡水刚毛藻类群不同于咸水刚毛藻和海洋刚毛藻类群,已经单独进化为一个支系。对中国内陆采集的69份刚毛藻样品进行统计分析后,Zhu等[11]提出目前淡水刚毛藻类群至少包含8个支系(clades)。由于各个支系的形态特征存在一定重叠性,这些支系暂未定种。Zhao等[17]在此基础上增加了淡水刚毛藻一新种硬刚毛藻Cladophora rigida,将淡水刚毛藻类群扩增到至少9个支系。
赤水河发源于云南省昭通市镇雄县,干流全长436.5 km,是长江上游南岸较大的一级支流。流域地貌以中山丘陵为主,景观类型多样,是长江上游为数不多的没有修建水坝和水库的支流,仍保持着天然的河流特征[18,19]。赤水河还是“长江上游珍稀、特有鱼类国家级自然保护区”的重要组成部分,因其生物多样性丰富,是长江上游一条独具价值的河流[20,21]。目前,有关赤水河流域藻类多样性的研究较少,且主要集中在对浮游植物群落结构的调查等方面[22,23]。本研究拟对冬季赤水河流域的刚毛藻进行广泛调查,以评估该流域刚毛藻多样性,为今后赤水河流域的生态环境保护提供基础数据和科学支持。
1 材料与方法
1.1 样品采集
本研究对冬季赤水河流域开展刚毛藻样品的广泛采集,在上、中、下游共设计样点38个 (图1)。使用镊子、刀片采集样品,并放置于采样瓶或采样袋中。采集到的丝状绿藻材料一部分保存于10%的福尔马林溶液中用于形态学观察,一部分保存于95%乙醇溶液中用于总DNA提取。环境样品分别保存于南宁师范大学植物学实验室和中国科学院水生生物研究所。采集样品带回实验室后通过Leica DM5000B显微镜(Leica,Wetzlar,Germany)进行镜检,筛选符合要求的样品以备下一步进行DNA提取。
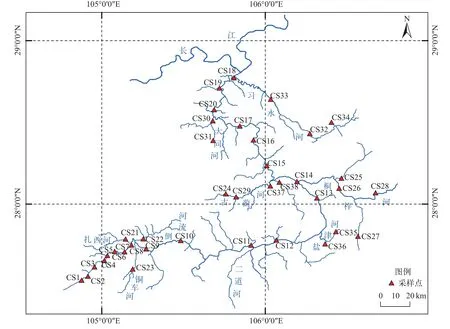
图1 赤水河流域采样点图Fig. 1 Location of sampling site at Chishui River basin
1.2 DNA提取及PCR扩增
在解剖镜下对目标样品进行小心分离,挑取干净的目的藻丝用ddH2O反复洗涤3—4次,在显微镜下镜检后置于含有磷酸盐缓冲液(PBS,pH=7.0)及玻璃破碎珠的2 mL冻存管中。使用mini-beadbeater(Model 3110BX,Biospec Products,Bartlesville,Oklahoma,USA)震荡破碎3min,重复3次直至藻丝破碎、内含物充分释放。最后用植物基因组提取试剂盒(Tiangen Biotech Co.,Beijing,China)对藻丝基因组总DNA进行提取,获得的总DNA保存于-20℃冰箱备用。
PCR扩增的反应体系: 2 μL模板DNA(10 ng/μL),1.25 UTaqDNA聚合酶(ExTaq,TaKaRa),正向引物和反向引物各0.2 μmol/L,0.4 mmol/L dNTPs和5 mmol/L MgCl2,加水补充至50 μL。SSU rDNA的序列由SR1-SS11H和SSU897-18SC2[6]两对引物扩增获得。PCR反应条件: 94℃预变性3min;94℃变性1min,55℃退火1min,72℃延伸1min 30s,此过程35个循环;最后72℃延伸3min[13]。LSU rDNA序列扩增使用的引物为C1 (F)和D2 (R)[8,24]。PCR反应条件: 94℃预变性5min;94℃ 30s,57℃ 30s,72℃30s,循环31次;72℃延伸5min。参考Hayakawa等[2]的引物Clado ITS-9F和Clado ITS-7R扩增内转录间隔区ITS序列。PCR反应条件: 94℃预变性1min;98℃变性10s,65℃退火30s,68℃延伸2min,此过程30个循环;最后72℃延伸10min。
1.3 系统发育分析
从GenBank数据库下载相关序列,结合本研究所采集样品的分子序列共同构建刚毛藻科的系统进化树。本研究共建立了2个数据集(datasets):dataset 1,是基于SSU和LSU两分子标记联合构建的刚毛藻科数据集(共80条序列,选取硬毛藻Chaetomorpha作为外类群);dataset 2,是基于SSU,ITS和LSU三分子标记联合构建的淡水刚毛藻数据集(共51条序列,选取咸水刚毛藻序列作为外类群)。使用MAFFT 7.0[25]进行序列比对,并在MEGA 7.0[26]中进行手工矫正。使用DAMBE 5.6[27]对比对变异位点进行突变饱和评估分析。通过PAUP 4.0b/MrModeltest v3.7[28]进行最佳模型测试。两个数据集均同时采用贝叶斯法(BI)和最大似然法(ML)进行系统进化分析。ML分析采用GTR+I+G模型在RAxML v8.1.2[29]中运行分析,用自展(Bootstrap,BP)分析检验,开展1000次随机重复抽样。贝叶斯分析采用MrBayes v.3.2[30]软件分析,运行1×106代,每1000代取样一次,分析达到平稳(标准误差<0.01)则运行结束。最大似然树和贝叶斯树分别在相应分支上获得自展支持值(BP)和后验概率值(PP)。采用MEGA 7.0[26]软件对所构建出的进化树各支系进行支系内和支系间的遗传距离计算。最后综合系统支持值、遗传距离和形态数据对淡水刚毛藻进行分类讨论。
2 结果
本研究共设计采样位点38个,获得的刚毛藻目样品(包含刚毛藻属、根枝藻属(Rhizoclonium)和黑孢藻属(Pithophora)样品)覆盖采样点21个,分布有刚毛藻属样品的位点19个,隶属于赤水河、倒流河、古蔺河、桐梓河、观音寺河、大同河和习水河,覆盖赤水河流域的上、中、下游(表1) (注: 二郎镇以上为上游,复兴镇则为中、下游分界点)。对照样品采集信息及系统发育结果,冬季赤水河流域刚毛藻数量和多样性分布整体呈现从上游向下游逐渐减少趋势。

表1 各采样点分布刚毛藻样品情况Tab. 1 The distribution of Cladophora samples at each sampling site
筛选符合要求的样品进行DNA提取,目标样品的生境、编号及GenBank号等信息见表2。对所有刚毛藻样品分别进行SSU、LSU和ITS三分子标记序列扩增,获得有效序列47条,其中同时成功获得三标记序列的有效样品数有14个。扩增所得的SSU、ITS和LSU序列长度约为: 1650、600和1000 bp。基于80条序列联合(Dataset 1,长约2550 bp)构建的SSU+LSU系统树主要展示赤水河刚毛藻藻株在刚毛藻科内的系统进化位置(图2)。该树包含了刚毛藻科主要的3个属: 刚毛藻属、根枝藻属和硬毛藻属,其中刚毛藻属和根枝藻属具有较近的亲缘关系。研究中所有采集自赤水河流域的刚毛藻均隶属于淡水刚毛藻支系(图2,BI=1.00;BP=95)。基于51条序列联合(Dataset 2,长约3183 bp)构建的SSU+ITS+LSU的系统树主要展示赤水河流域刚毛藻在刚毛藻属内所处的系统进化位置(图3)。基于最大似然法构建的系统进化结果和基于贝叶斯法构建的系统树结果基本一致,图示(图2和图3)仅展示贝叶斯建树结果的拓扑结构。

表2 本研究所用刚毛藻样品的采集信息及序列信息Tab. 2 Cladophora specimens used in this study,and associated collection information and GenBank accession numbers
基于核糖体序列所构建的系统发育结果(图2和图3)与Zhu等[11]及Zhao等[17]的研究结果基本一致。为保持研究的持续性及便于对比分析,本研究对淡水刚毛藻各clade的划分和标注与前述文献[11,17]保持一致。加入赤水河样品后,本研究将淡水刚毛藻类群由此前描述的“至少包含9个支系”扩展到“至少包含10个支系”。新增的淡水支系clade 10(BI=1.00;BP=100)当前仅包含两条序列(CS0801CS2202),藻株分别采自赤水河和倒流河,系统树图显示clade 10和clade 7具有较近的亲缘关系(BI=1.00;BP=100;图3)。基于SSU+LSU双分子标记和基于SSU+ITS+LSU三分子标记的系统进化结果基本一致,部分支系间的亲缘关系和支持值有所差异。加入ITS序列所构建的进化树,除clade 4支持度不高外,其他支系支持度良好(BI=1.00;BP>95;图3)。基于三分子标记的进化结果可将淡水刚毛藻类群划分为两组(group): 其中clade 1、clade 2和clade 4三个支系具有较近的亲缘关系,聚为一组(Group Ⅰ;BI=1.00;BP=94);clade 5、clade 6、clade 7、clade 9和clade10五个支系具有较近的亲缘关系,聚为另一组(GroupⅡ;BI=1.00;BP=100;图3)。系统发育分析结果显示,赤水河流域采集的样品分属于6个支系,覆盖淡水刚毛藻类群支系(10个支系)一半以上。既有隶属于Group Ⅰ的: clade 1 (CS0901CS3401CS2501)、clade 2 (CS1401)和clade 4 (CS3202),也有隶属于Group Ⅱ的clade 7 (CS1101CS1102CS2401)、clade 9 (CS0821CS3003CS3003YN1901)和clade 10 (CS0801CS2202)。各支系间和支系内的遗传距离计算结果见表3和表4。从结果看,各支系内部的最大遗传距离均≤0.003,表明各支系所含序列遗传差异较小(表4)。各支系间的遗传距离变化范围大(0.004—0.023),其中clade 8和其他支系间的遗传距离均较大(0.019—0.023),新划分的clade 10和clade 7之间的遗传距离为0.006,该值和clade 5和clade 6之间的遗传距离相同,高于clade 1和clade 4(0.004)、clade 2和clade 4 (0.005)间的遗传距离(表3)。

图2 基于SSU+LSU 两个分子标记构建的刚毛藻科系统进化树Fig. 2 Bayesian SSU+LSU (based on dataset 1) phylogeny of Cladophoraceae
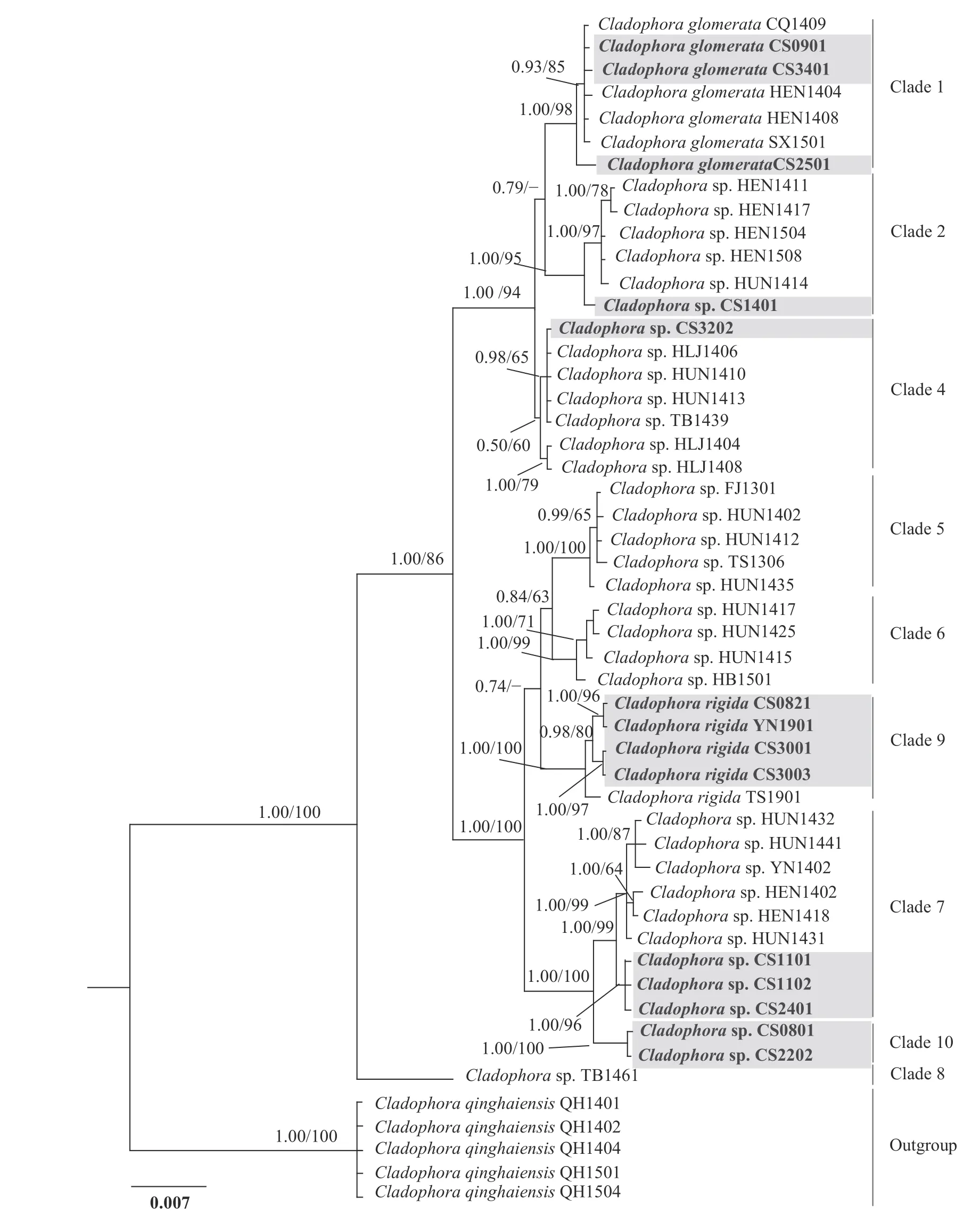
图3 基于SSU+ITS+LSU 三个分子标记构建的淡水刚毛藻系统进化树Fig. 3 Bayesian SSU+ITS+LSU (based on dataset 2) phylogeny of the freshwater Cladophora
3 讨论
由于刚毛藻形态上的简单性、可塑性(易受环境条件影响),使得单独依靠传统分类特征(如细胞直径、分枝类型、生长方式和固着器)进行刚毛藻种类鉴定十分困难。本研究首次大规模地对赤水河流域的刚毛藻展开调查,并基于多分子标记进化系统发育分析。冬季赤水河流域刚毛藻多样性较高,所采藻株分属于6个支系,覆盖当前淡水刚毛藻支系一半以上。此外,刚毛藻在该流域分布广泛,上、中、下游均有分布(表1)。其中,上游分布最多(11个位点),中游其次(5个位点),下游较少(3个位点),这可能与上、中、下游水质状况、透明度及水体流速等存在一定关系。
核糖体分子标记是刚毛藻科系统分类研究中常用的分子标记,常单独使用或者联合使用[11,31]。本研究分别基于SSU+LSU双分子标记和SSU+ITS+LSU三分子标记构建了刚毛藻属的系统进化树。结果显示,加入ITS分子标记后,淡水刚毛藻内部各个支系间的亲缘关系更加明确、支持强度和聚类结果更好。例如: clade 1、clade 2、clade 5和clade 6的这几个支系的自展支持值(BP)从小于75升至95以上。表明较为保守的分子标记(SSU和LSU rDNA)的联合适于属以上水平的进化分析,联合变异速率较快的分子标记(如ITS序列)则有利于属以下、种水平的进化分析,这一结论也在近年来刚毛藻科相关的系统学研究中得到支持[11,17]。
有关刚毛藻不同支系的划分目前尚无严格的标准,本研究主要参考已有的文献研究结果,同时结合不同支系间的遗传距离及部分形态特征来划分。例如,基于SSU+ITS+LSU三分子标记联合分析,Zhu等[11]将以HLJ1406为代表的藻株归入clade 4分支,该支系支持值不高(-/50)。在本研究中,加入了赤水河流域刚毛藻序列后,clade 4的系统支持值有所提升,但提升也不明显(0.50/60)。形态上,该分支内部分藻株形态和C. rivularis(Linnaeus) Hoek 1963相似: 如多营漂浮生长,呈现弱化的向顶生长,藻丝长、分枝稀疏。但也有研究将采自中国的样品(编号P02,鉴定为C.cf.glomerata)和采自冰岛的样品(编号B50,鉴定为球刚毛藻C. globulina)归入clade 4。该支系的藻类究竟是溪生刚毛藻C. rivularis或球刚毛藻C. globulina还是应归入种以下单元目前尚不能确定。赤水河藻株CS1401和clade 2内其他序列虽然具有一定碱基差异,但将其划分至clade 2时,该分支内的平均遗传距离仅为0.001,最大遗传距离值0.003,低于其余分支间的遗传距离(表3和表4)。在形态上,CS1401藻株幼小时基部区域分枝较多,上端分枝少;成熟藻丝很长,可平行排列、相互缠绕;具有固着器。这些形态和Zhu等[11]所描述的clade 2形态具有相似性,因此将CS1401划分至clade 2。此外,值得关注的藻株是CS0801和CS2202,二者形成的新支系clade 10和clade 7具有较近的亲缘关系。形态上CS0801、CS2202也和clade 7藻株具有部分相似性: 均具有基部固着器,细胞直径大小相近,CS0801有和clade 7藻株类似的分支卷曲现象,CS2202有和clade 7藻株类似的分枝偏侧生现象。但是CS0801和CS2202藻株也具有不同于clade 7的一些特征,如藻丝手感细软(细胞有时因其柔软而弯曲或变形),细胞居间分裂占优势,CS0801藻株一个细胞上常可形成3—4个细胞轮生的现象。在遗传距离上,当clade 7不包含CS0801、CS2202序列时,分支内平均遗传距离为0.002,最大的遗传距离为0.003,低于其余分支间的遗传距离。加入CS0801、CS2202后,clade7分支内平均遗传距离为0.003,最大的遗传距离为0.007,该值高于clade 1与clade 4,clade 2与clade 4,clade 5与clade 6这3组分支间的遗传距离,表明CS0801、CS2202和clade 7内其他藻株序列差异较大(表3和表4)。综合形态和序列数据,本研究将CS0801、CS2202划分至一个新clade,即clade10。
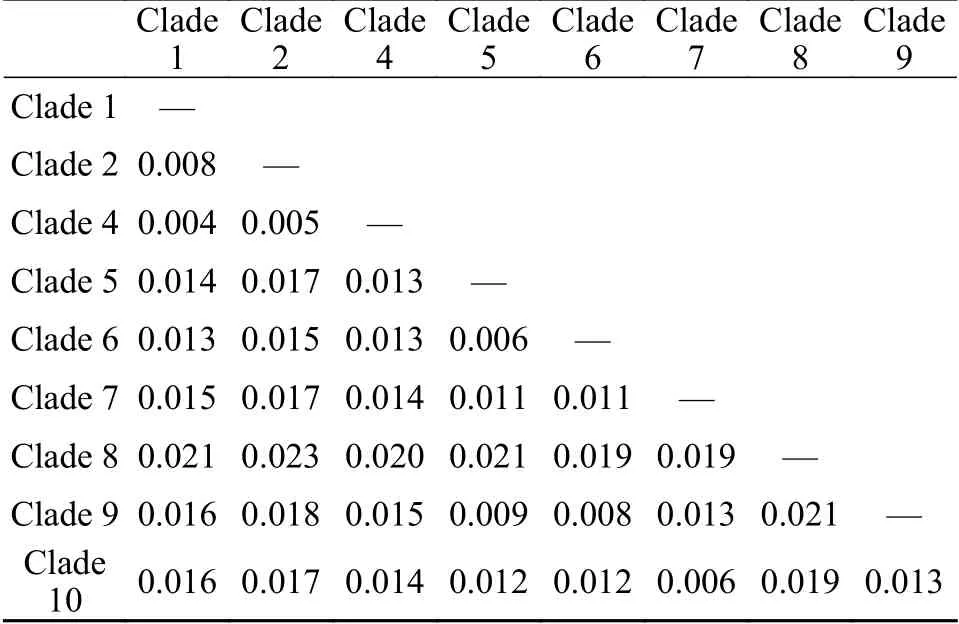
表3 淡水刚毛藻各支系间的遗传距离(基于SSU+ITS+LSU)Tab. 3 Between clade of freshwater Cladophora mean distance(based on SSU+ITS+LSU)
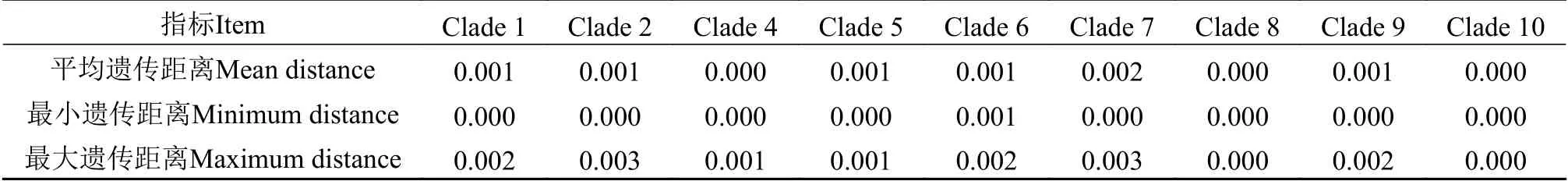
表4 淡水刚毛藻各支系内的遗传距离(基于SSU+ITS+LSU)Tab. 4 Within clade of freshwater Cladophora distance (based on SSU+ITS+LSU)
尽管本研究未进行详细的形态学报道,该流域所采集的4株编号为CS0821、CS3001、CS3003和YN1901 (clade 9)的刚毛藻藻株仍具有不同于典型刚毛藻的一些形态特征,如: 藻体不分枝,细长、簇状,手感较硬,细胞圆柱形,直径较大(180—429 μm),长宽比较小(0.5—3.3),通过固着器或者基部假根着生于水中岩石表面,同时可以产生丰富的侧生假根。基于该种独特的形态特征及分子数据支持,前期研究将其确定为刚毛藻属一新种: 硬刚毛藻C. rigida[17]。该种较为少见,目前仅在赤水河流域和湖北咸宁有报道。除了clade 9藻株具有典型形态外,clade 1和clade 8也具有较为典型的特征。clade 1藻丝分枝十分旺盛、藻丝多呈鲜绿色,具有显著的顶端生长优势,将其确定为团集刚毛藻C. glomerata。Clade 8藻株目前仅报道于西藏地区,该支系位于淡水刚毛藻类群的基部,和青海刚毛藻(咸水种)有较近亲缘关系[11,32]。Clade 8藻丝非常纤细,顶端细胞直径大小仅为19.8—30.5 μm,顶端细胞末端会呈现典型的锥状或刺状;分枝有时呈“Z”字形,与主枝夹角成钝角[11]。当前,一些典型刚毛藻种类如: 溪生刚毛藻C. rivularis、脆弱刚毛藻C. fracta和球刚毛藻C. globulina等,缺乏完整有效的分子数据,并未加入到SSU+ITS+LSU联合分析中。模式标本缺乏有效的分子序列、不同支系间形态特征的重叠性及缺乏标准的诊断特征都使得当前对各个clade给出确切种名较为困难。
值得注意的是,近年来随着全球越来越多刚毛藻目样品的获取和分析,传统依靠形态特征所定义的刚毛藻目内各属的界限正逐渐被打破: 如分枝根枝藻(R. ramosum)和厚壁根枝藻(R. pachydermum)具有类似刚毛藻的真正分枝;本研究采集的硬刚毛藻(C. rigida)展现的典型的不分枝形态、较大的细胞直径和较小的细胞长宽比使其看起来更像硬毛藻;中国发现的10亿年前的海藻化石Proterocladus antiquus,其形态则和扩展刚毛藻C. herpestica、分枝根枝藻R. ramosum非常相似[17,33—35]。这种不同属间形态特征的相似性和多样性形成原因尚不清楚。有研究显示,石莼纲是绿藻门中形态和细胞多样性极其丰富的一个纲,刚毛藻目又是该纲代表性的一大类群,石莼纲内藻类的多样性可能与该纲分化自成冰纪(Cryogenian)或之前时期所处的特殊地球环境有关[35,36]。
本研究对赤水河流域刚毛藻多样性开展了调查,采集生境以河流沿岸为主。实际上刚毛藻具有十分广泛的地理分布,除河流沿岸带以外,湖泊、水库、池塘、沟渠、瀑布、潮湿地表甚至树皮等亚气生环境也适于其生长。更广泛生境的样品调查将是下一步研究的内容,也将帮助我们进一步理解刚毛藻目样品的地理分布及其演化关系。
