碳氮源对新古尼异虫草抗氧化活性的影响
2022-08-19林群英龚光禄龙良鲲钱奎宪邓春英
林群英,龚光禄,龙良鲲,钱奎宪,邓春英
1. 中华全国供销合作总社南京野生植物综合利用研究所(南京 211100);2. 贵州省农业科学院,贵州省食用菌育种重点实验室(贵阳 550006);3. 南京林业大学化学工程学院制药工程系(南京 210037);4. 贵州科学院(贵阳 550001)
新古尼异虫草(Metacordyceps neogunnii)是一种隶属于麦角菌科(Clavicipitaceae)的大型真菌,大量分布于我国的贵州、湖南、云南等地,一直以来被视为古尼虫草(Cordyceps gunnii),直到2017年,才被证实为新种,命名为新古尼异虫草[1]。这一分类学研究成果为新古尼异虫草的研究提供更准确的分类定位,突显其研究的重要性。
新古尼异虫草这一资源在我国民间应用历史悠久,在贵州等产地甚至被作为冬虫夏草的替代品,功效价值备受肯定[2-3]。生物活性成分研究证实,新古尼异虫草含有黄酮、多酚、多糖等多种与抗氧化密切相关的活性物质,被证实具有抗氧化、抗肿瘤、镇痛、镇静和免疫调节等作用[4]。抗氧化是虫草资源重要的生物活性[5],但新古尼异虫草在这方面的研究相对较少,其菌丝体可作为维生素E制备的原料[6],经酶解的胞内多糖具有抗氧化活性[7]。张永明等[8]以野生子实体为材料提取总黄酮,但未见以培养产物为提取材料的研究。
新古尼异虫草的人工培养可追溯至1985年,通过组织分离方式获得纯培养物[9]。新古尼异虫草在人工培养方面已取得一定进展。曹蕾等[10]筛选得其菌丝体液体培养的最佳碳氮源,分别是甘露糖和尿素;刘晓翠等[11]则认为蔗糖和蛋白胨分别为最佳的碳氮源。由于这些研究结果不一致,有必要对新古尼异虫草的培养进行研究。培养条件对这种虫草的菌丝体抗氧化活性及活性物质含量的影响未见报道。通过液体培养条件的调控可获得特定产物,且易于分离。应用这种方式,有针对性地获得富含黄酮和多酚的菌丝体,是开发新古尼异虫草抗氧化产品的有效途径。试验以自行分离的新古尼异虫草菌株为材料,系统分析该真菌积累黄酮和多酚等抗氧化活性物质的营养条件,利用体外试验评价产物的抗氧化能力,为抗氧化产品的开发提供理论依据。
1 材料与方法
1.1 供试菌种
野生新古尼异虫草子实体,采集自贵州省施秉县。虫体结实饱满,子座幼嫩未成熟(图1)。

图1 野生新古尼异虫草
1.2 菌种分离及真实性鉴定
1.2.1 菌种分离
野生新古尼异虫草带回实验室进行菌种分离。清除虫体表面泥沙等杂物及表面网丝状物,用75%酒精棉擦拭材料。采用组织分离的方式进行菌种分离,用无菌镊子夹取子实体和虫体菌核中的菌肉,接种至PPDA斜面培养基中,在25 ℃下培养14 d。菌种经转管纯化后用于培养。
1.2.2 分子鉴定
经液体培养获得新古尼异虫草菌丝体,用无菌水冲洗4次,沥干水分,用无菌滤纸尽量吸干菌丝体的水分,用于DNA提取。采用CBTA法提取总DNA,利用通用引物进行ITS序列扩增,由上海生工公司完成测序工作。将序列上传至GenBank获得序列号,选取已发表的相似性高的序列,采用邻接法进行系统树构建,进一步明确其分类地位,完成菌种真实性的分子鉴定。
1.2.3 无性型形态观察
将直径5 mm的菌块接种至PPDA平板培养基中央,用封口膜封好。置于25 ℃进行暗培养,培养14 d,观察菌落特征,包括气生菌丝、色素形成、菌丝致密程度等。采用电子扫描显微镜观察菌丝体和分生孢子形态特征。
1.3 碳源和氮源对新古尼异虫草菌丝体生长的影响
将6种碳源分别添加至空白培养基(蛋白胨4 g/L、牛肉浸膏4 g/L、KH2PO41.0 g/L、MgSO4·7H2O 0.5 g/L、维生素B10.05 g/L、pH 6.5、CK-C)中,用量20 g/L,配制液体培养基。按5%(V/V)接种量接入一级液体菌种,按25 ℃、160 r/min振荡培养6 d,中止培养。用约0.104 mm(150目)孔径尼龙布收集菌丝体,菌丝体用水冲洗3次以上,于60 ℃烘干至恒重,称其质量。以空白培养基(CK-C)为对照,各组设置3个平行。将6种氮源分别添加至空白培养基(葡萄糖20 g/L、KH2PO41.0 g/L、MgSO4·7H2O 0.5 g/L、维生素B10.05 g/L、pH 6.5、CK-N)中,用量8 g/L,配制液体培养基。余下步骤同前。以空白培养基(CKN)为对照,各组设置3个平行。
1.4 抗氧化活性评价
1.4.1 提取物的制备
将1.3小节中各组培养所得的菌丝体研磨成0.85 mm(20目)粒径大小的粉末,精确称取0.2 g,按照料液比1∶10 g/mL加入80%乙醇,在80 ℃下水浴加热提取1 h,按10 000 r/min离心10 min,取上清液,共提取2次,合并上清液,定容至10 mL,即为醇提物。剩余残渣挥尽乙醇,按照料液比1∶10 g/mL加入蒸馏水,沸水浴提取1 h,按10 000 r/min离心10 min,取上清液,共提取2次,合并上清液,定容至10 mL,即为水提物。
1.4.2 总黄酮及总多酚含量测定
总黄酮含量采用AlCl3-NaNO2方法测定[12];总多酚含量采用酒石酸铁法测定[13]。
1.4.3 DPPH·清除率测定DPPH·清除率的测定参照叶琦等[14]的方法进行,取各提取物100 μL用于测定。
1.5 数据分析
采用Excel 2010和SPSS 18.0进行数据分析和作图。
2 结果与分析
2.1 菌种真实性鉴定
2.1.1 分子鉴定
分离获得一株新古尼异虫草菌种,编号为Cg7,保存于南京野生植物综合利用研究所。该菌株ITS序列全长631 bp,序列号为MZ725544。根据系统进化树的构建结果,MZ725544与Wen等[1]报道的新古尼异虫草的序列KU729718.1和KU729719.1等聚为一个分支(图2),证实试验所分离所得的菌株Cg7是新古尼异虫草(Metacordyceps neogunnii)。
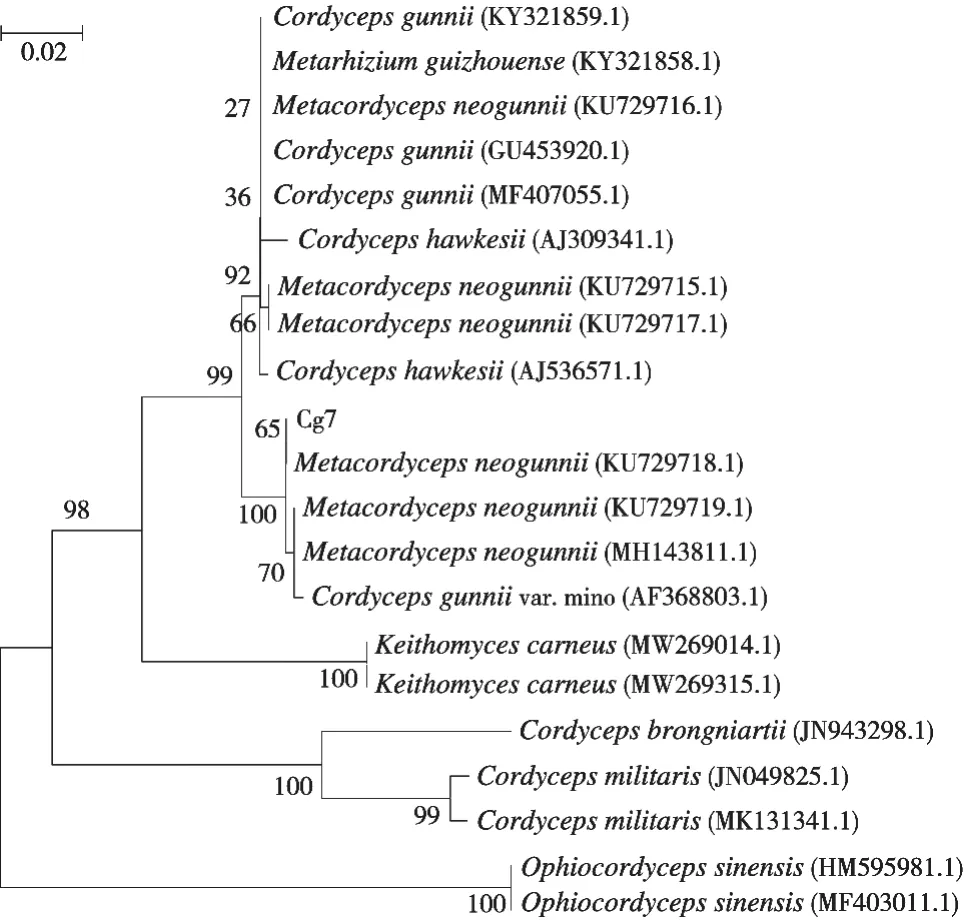
图2 基于ITS序列构建的新古尼异虫草系统进化树(邻接法)
2.1.2 无性型形态观察
新古尼异虫草菌株Cg7生长14 d,菌落直径5.6 cm,呈致密绒毛状,具有较明显的轮层,背面呈墨绿色(图3 a和b)。通过扫描电子显微镜观察到新古尼异虫草分生孢子具有明显的刺状物(图3 c)。
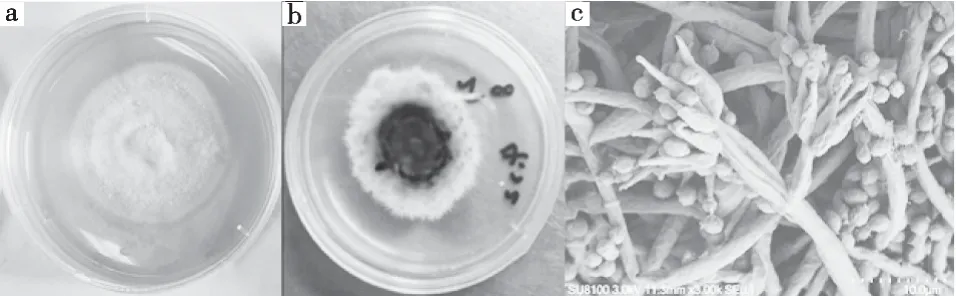
图3 新古尼异虫草菌落形态及产孢结构特征
2.2 碳氮源对新古尼异虫草菌丝体产量的影响
新古尼异虫草对部分小分子碳源具有较好的利用能力,对甘油、葡萄糖和蔗糖的利用效果十分接近,无显著性差异,菌丝体产量分别为11.45,10.67和11.50 g/L,其次是麦芽糖,产量为10.33 g/L,与空白对照组CK-C有显著性差异(图4)。乳糖和可溶性淀粉利用能力较差,菌丝体有一定的生长,产量约为2 g/L。所有测试的6种碳源与CK-C均有显著性差异,不添加碳源的菌丝几乎不生长。可见,菌丝体培养可选用甘油、葡萄糖或蔗糖。
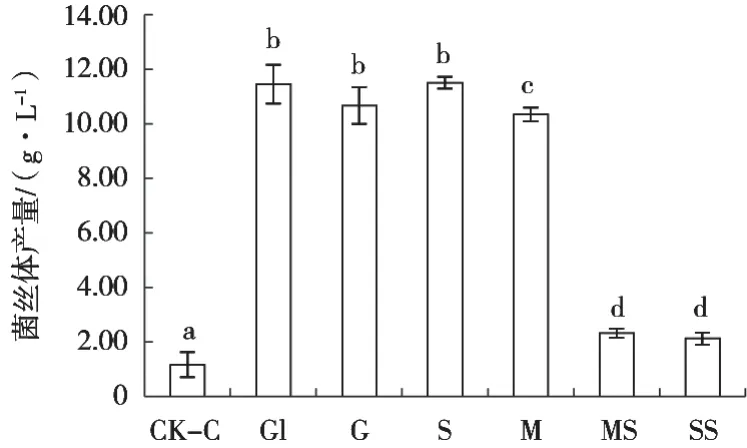
图4 碳源对新古尼异虫草菌丝体产量的影响
6种有机氮源对菌丝体产量的促进作用均十分明显,其中蛋白胨的效果最好,达13.11 g/L,而牛肉浸粉的效果相最差,但菌丝体产量在9 g/L以上,远高于不添加氮源的空白对照组(CK-N,图5)。在缺乏氮源的条件下,新古尼异虫草的菌丝体仅有少量生长,产量只有1.22 g/L。因此,蛋白胨是新古尼异虫草菌丝体生长的最佳氮源。
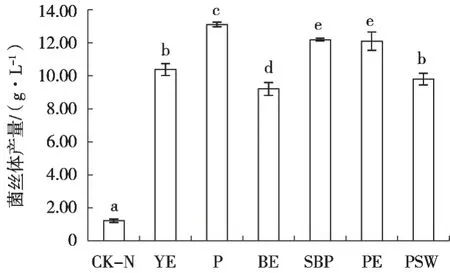
图5 氮源对新古尼异虫草菌丝体产量的影响
2.3 碳氮源对新古尼异虫草菌丝体中总黄酮含量的影响
在各碳源培养所得的菌丝体中,醇提物的总黄酮含量均高于水提物。可溶性淀粉作为碳源培养获得的菌丝体中总黄酮含量最高,达4.9 mg/g,其次是乳糖,为2.8 mg/g(图6)。而最利于菌丝体生长的蔗糖对总黄酮的生成促进作用最小,含量仅为1.2 mg/g。利于菌丝生长的甘油和葡萄糖对总黄酮的积累有较好的促进作用,含量均为2.5 mg/g。由于可溶性淀粉为碳源时菌丝体的产量仅约为葡萄糖或甘油的20%,因此,结合菌丝体生物量及总黄酮含量,葡萄糖和甘油均可作为富含黄酮菌丝体的碳源。
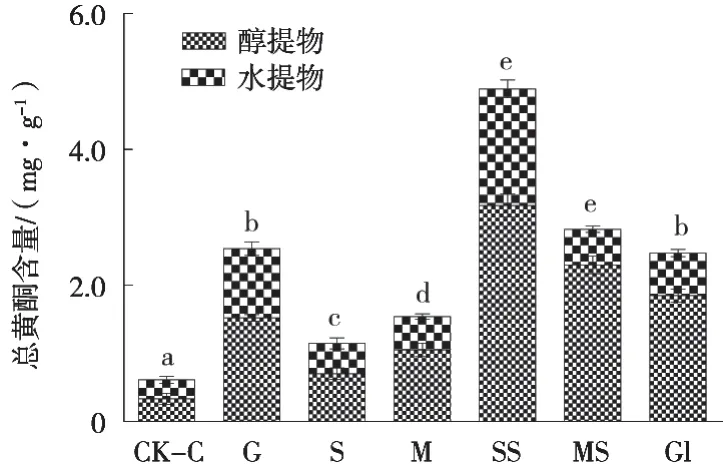
图6 碳源对新古尼异虫草菌丝体中总黄酮含量的影响
6种有机氮源均能有效地促进新古尼异虫草中总黄酮的形成。酵母浸出粉是新古尼异虫草菌丝体积累黄酮的最佳氮源,总含量达3.9 mg/g,其中醇提物和水提物中的含量分别为2.3 mg/g和1.6 mg/g,其次是牛肉浸粉,总含量为3.5 mg/g,黄豆粉次之(图7)。而土豆浸出粉对总黄酮的形成影响最小,含量为2.6 mg/g,比空白对照组高了近2倍。结合菌丝体生物量考虑,选择黄豆粉为新古尼异虫草形成总黄酮的最佳氮源。

图7 氮源对新古尼异虫草菌丝体中总黄酮含量的影响
2.4 碳氮源对新古尼异虫草菌丝体中总多酚含量的影响
在各碳源培养所得的新古尼异虫草菌丝体中,醇提取中总多酚的含量均高于水提物。最利于总多酚积累的碳源是蔗糖,总含量为16.8 mg/g,其中醇提物的含量为14.4 mg/g,其次是麦芽糖,总含量为14.5 mg/g(图8)。而利于菌丝生长的甘油则对总多酚的积累无显著的促进作用,仅为3.4 mg/g,仅比空白对照组高了0.3 mg/g,无显著性差异。蔗糖对菌丝体的产量和总多酚的积累均为最佳,因而是新古尼异虫草生产总多酚的最佳碳源。氮源中蛋白胨对新古尼异虫草菌丝体中总多酚含量影响最显著,含量为9.7 mg/g,其中醇提物中含量最高,为6.5 mg/g,其次是黄豆粉。而酵母浸出粉为氮源时,水提物的总多酚含量最高,为4.6 mg/g(图9)。结合菌丝体生物量因素,蛋白胨是新古尼异虫草产生总多酚的最佳氮源。

图8 碳源对新古尼虫草菌丝体中总多酚含量的影响

图9 氮源对新古尼虫草菌丝体中总多酚含量的影响
2.5 碳氮源对新古尼虫草菌丝体DPPH·清除能力的影响
葡萄糖为碳源时,菌丝体的醇提物对DPPH·清除率最高,达92.74%,其次是可溶性淀粉和麦芽糖,分别为85.18%和84.34%(图10)。其余3种碳源培养的菌丝体的提取物均低于空白对照组,未能提高菌丝体的抗氧化活性。而水提物方面,只有可溶性淀粉对菌丝体的抗氧化活性具有促进作用,而甘油作碳源的菌丝体水提物清除率最低,仅为18.20%。
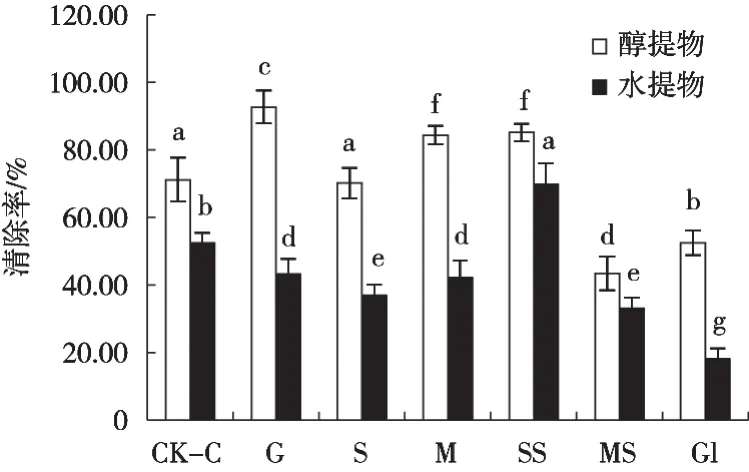
图10 碳源对新古尼虫草菌丝体抗氧化活性的影响
氮源对菌丝体的抗氧化活性有较好的促进作用。牛肉浸粉、蛋白胨和蚕蛹粉培养所得的菌丝体的醇提物对DPPH·清除率均高于空白对照组,其中以牛肉浸粉最高,为91.51%(图11)。仅蚕蛹粉和黄豆粉为氮源获得的菌丝体水提物的清除率高于空白对照组,且高于其醇提物的清除率。土豆浸出物为氮源获得的菌丝体水提物对DPPH·的清除率最低,仅为17.11%。

图11 氮源对新古尼虫草菌丝体抗氧化活性的影响
3 讨论与结论
新古尼异虫草是以昆虫为寄主的真菌,其内菌核定殖了其他微生物[15],加之该虫草子实体纤细,导致所分离菌种纯度易受影响,分离难度比一般大型真菌大。从其他虫草资源的研究历程来看,菌种的真实性鉴定是有必要的。因此,试验对所采用的菌株进行ITS测序和系统树构建,结合无性型的形态特征,明确该菌株的真实性,为后续研究提供真实有效的新古尼异虫草菌种。
新古尼异虫草的菌株差异对研究有重要影响。根据蒋岚等[16]的研究,新古尼异虫草菌株可划分为3个类型,其中产棕色色素的菌株菌丝体生物量最高,活性成分含量高,适用于规模化生产。而贾学渊等[17]则认为产绿色素的菌株产活性成分能力更强。试验采用的菌株Cg7属于产绿色素的类型(图3b)。鉴于菌株类型与活性之间未有确切的相关性,以自行分离的菌株为材料,对影响菌丝体生长、黄酮和多酚等次生代谢成分的积累及抗氧化活性的碳氮源情况进行较系统的分析。菌株Cg7菌丝体生长的最佳碳氮源分别是蔗糖和蛋白胨,与曹蕾等[10]和刘晓翠等[11]的报道不完全一致。菌株Cg7对可溶性淀粉的利用能力极差,产量仅为3 g/L,远低于刘晓翠等[11]报道的14.7 g/L。因此,对于新获得的菌株,在进行规模化应用前有必要对其进行系统的生物学研究。
新古尼异虫草含有多种活性物质,包括黄酮和多酚[3,8],多糖也被证实具有较好的抗氧化活性[7],其抗肿瘤活性受培养基的影响[18]。为了更有针对性地提高培养产物的产量,对新古尼异虫草菌丝体生长及与抗氧化相关的黄酮和多酚积累条件进行初步筛选。最利于黄酮积累的碳源和氮源分别是可溶性淀粉和酵母浸出物,其含量分别为4.9和3.9 mg/g,远高于九州虫草总黄酮的含量1.89 mg/g[19]。筛选获得最利于新古尼异虫草菌丝体多酚积累的碳氮源,即蔗糖和蛋白胨,总含量分别为16.8和9.7 mg/g,高于口蘑等食用菌中的含量[20]。
不少试验证实,多酚与抗氧化活性存在较明确的相关性,可有针对性地培养生产。从新古尼异虫草菌丝生长情况可得知,这个菌株对碳氮源有着较严格的要求,除了具有明显的选择性外,在缺乏碳源或氮源时,其生长均受到显著的限制,继而影响次生代谢产物黄酮和多酚的产量,但菌丝体的生物量与次生代谢产物含量无相关性,这与多糖发酵的结果一致[11]。
利用体外清除DPPH自由基的方法测定新古尼异虫草不同发酵产物的抗氧化活性,并测定与抗氧化密切相关的黄酮和多酚的含量,研究发现它们之间无相关性,这与已有报道相一致[21]。根据章能胜等[22]的试验结果,从古尼拟青霉中分离纯化得到2个高纯度的强抗氧化活性成分,可推测菌丝体中可能含有其他可清除DPPH·的物质。通过此研究证实新古尼异虫草是抗氧化活性研究的潜在大型真菌,为其开发应用提供有力参考依据。
