斑石鲷消化系统形态学与组织学观察
2022-08-16肖志忠肖永双姬广磊邓钦有王雨福马玉婷刘相全连亚明杨理忠
陈 宵, 肖志忠, 肖永双, 姬广磊, 邓钦有, 李 旋, 王雨福, 马玉婷, , 6, 李 军, 刘相全, 连 昌, 连亚明, 杨理忠
斑石鲷消化系统形态学与组织学观察
陈 宵1, 2, 3, 4, 肖志忠1, 8, 9, 肖永双1, 4, 姬广磊5, 邓钦有2, 3, 李 旋2, 3, 王雨福1, 4, 马玉婷1, 4, 5, 6, 李 军1, 4, 刘相全2, 连 昌7, 连亚明8, 杨理忠9
(1. 中国科学院 海洋研究所/中国科学院 海洋大科学研究中心 山东省实验海洋生物学重点实验室, 山东 青岛 266071; 2. 山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室, 山东 烟台 264006; 3. 上海海洋大学 水产与生命学院, 上海 201306; 4. 青岛海洋科学与技术试点国家实验室 海洋生物学与生物技术功能实验室, 山东 青岛 266237; 5. 威海市海洋发展研究院, 山东 威海 264200; 6. 青岛农业大学 海洋科学与工程学院, 山东 青岛 266109; 7. 威海市文登区海和水产育苗有限公司, 山东 威海 264400; 8. 威海市好慧淦海洋生物科技有限公司, 山东 威海 264400; 9. 莆田市蒲盛水产科技有限公司, 福建 莆田 351174)
为探明斑石鲷()消化系统的形态特征和组织学结构, 助力斑石鲷的人工繁育及养殖技术研发, 作者采用形态解剖学和组织学切片技术对斑石鲷消化系统进行了实验研究。研究发现斑石鲷的消化道共由6部分组成, 从头至尾依次是口咽腔、食道、胃、幽门盲囊、小肠、直肠。斑石鲷的口咽腔内部空间大, 颌齿内部彼此连接、外部愈合, 呈现典型的鹦鹉喙状, 咽齿呈扁平的圆盾状, 于上下颌齿内侧上下对称分布, 这样复合结构(喙状颌齿与圆盾状咽齿)使得斑石鲷能够轻易地碾碎甲壳动物的外壳, 进而完成摄食。斑石鲷的食道短粗, 黏膜层密布具纵褶, 内部含有丰富的杯状细胞, 能够协助斑石鲷吞咽食物。胃部呈典型的不对称V型, 黏膜层下分布着密集的胃腺, 幽门部拥有整个消化道最厚实的肌肉层, 摄取的食物将在这里完成消化和分解。斑石鲷的小肠分为前肠、中肠、后肠3部分, 管径大小逐次递减, 黏膜层上肠绒毛的密度和长度也遵同样的规律, 从结构可以推断出斑石鲷肠道吸收功能主要集中于前中肠。直肠管径与小肠前肠段接近, 长度很短仅为小肠的1/7, 相比小肠部分, 肠绒毛更加稀疏, 绒毛长度也更短, 主要承担消化残留物的压缩排泄和水分与微量元素的重吸收, 相对简单结构对应着相对简单的功能。
斑石鲷(); 消化道; 形态学; 组织学
斑石鲷(), 隶属于鲈形目(Perciformes)、石鲷科(Oplegnathidae)、石雕属(), 俗称黑金鼓、斑鲷, 主要分布于日本、韩国、中国沿海以及菲律宾海域, 属于近海温水性鱼类[1]。斑石鲷肉质细嫩, 口味鲜美, 是制作生鱼片的绝佳食材, 深受国内和日韩高端海鲜餐饮市场的喜爱; 此外, 斑石鲷体型扁宽, 灰褐色的体表分布有均匀的黑色斑点, 极具观赏价值; 自2014年成功实现人工繁殖后, 已成为中国海水网箱和工厂化养殖优质经济鱼种, 同时也是海洋牧场增殖放流理想鱼种[2]。
消化系统对外界物质营养吸收和转化能力的强弱, 对鱼类的生长、发育和繁殖有着直接的影响。对消化系统进行形态学和组织学研究能够有效探明鱼类消化系统和摄食、消化、吸收之间的基础关系[3, 4]。国内外与此相关的研究报道颇多,如杨晓鸽等[5]对长麦穗鱼()的消化系统进行研究时发现, 其没有明显的胃组织, 肠道较短, 结合口裂特征, 判定其为杂食性; 胡玲玲等[6]则对条石鲷()的消化道形态学和组织学的研究过程中, 发现条石鲷食道和胃拥有4层组织结构, 而小肠部分则只有3层组织结构; 同样曹潇等[7]在黑鱾()的消化系统的形态学和组织学研究上则发现黑鱾吸收营养的主要场所是小肠的前、中肠部分。王雨福等[8]在2015年对斑石鲷早期发育特征进行了研究, 其中也涉及了消化系统在发育过程中的特点。本文重点为斑石鲷消化系统形态学和组织学上的观察和研究, 旨在对斑石鲷的生物学资料进行补充, 为后来者在斑石鲷的研究中提供一定的参考, 向斑石鲷繁育养殖方法的改善提供一些依据。
1 材料与方法
1.1 材料
解剖用斑石鲷于2021年7月取自山东文登海和育苗场, 共3尾, 平均体长26 cm, 体质量498.04 g, 样本活跃健康, 体表无明显病、伤痕迹。
1.2 方法
选取的斑石鲷转移至边长60 cm的玻璃水箱中暂养, 维持每天1/2的换水量及充分的换气, 期间停止投饵, 每12 h清理1次缸底的鱼粪, 约36 h后缸底不再出现明显的鱼粪, 判定已完成肠道排空, 对样本进行解剖。打开腹腔后将内脏完整取出, 从中分离出完整消化系统。观察消化系统各部分的形态特征并做记录。取食道、胃、小肠、直肠、肝脏、胰脏组织用Davidson’s固定液进行组织学固定; 使用全自动组织脱水机(Tp1020)完成样品的脱水和渗蜡; 使用石蜡包埋机(Histocentre2)对组织进行包埋处理; 待蜡块成型, 对其进行修整, 使用转轮式切片机(Leica rm2255)进行切片, 切片厚度为4~6 µm。切片烘干后进行HE染色, 最后用中性树胶封片, 使用显微镜(Olympus-CX31)进行观察、拍照。
1.3 数据采集和处理
小肠前中后肠绒毛的高度为绒毛顶端至基部凹陷处的垂直距离[9]; 宽度为单根肠绒毛最宽与最窄处的均值; 数量为相同倍率视野下, 3份组织样品在视野中最大绒毛数量的均值。利用GraphPad Prism 8.0和Spss v26.0对数据进行处理和分析。
2 结果
2.1 斑石鲷消化道形态特征
斑石鲷的消化道主要由6部分组成(图1), 分别为口咽腔、食道、胃、幽门盲囊、小肠、直肠。口咽腔内空间较大, 颌齿已愈合成一体, 外形呈鹦鹉喙状; 坚硬的臼齿于颌齿内对称分布, 臼齿圆盾扁平; 舌头呈椭圆形, 鳃弓于中轴线两侧对症分布, 每根鳃弓上存在两行梳状棘刺, 靠外侧端的棘刺较内侧更长。食道位于口咽腔后部, 形态短粗, 内表面密布纵褶, 具有极佳的扩张性。胃部是整个消化道最为膨大的部分, 外形呈不对称的V型, 以胃体部为分界点, 贲门侧胃腔的长度更长也更粗, 幽门侧则稍短也相对更细。幽门盲囊环绕附着于幽门和小肠的交界处, 整体呈碗状依附于胃幽门处, 以肠轴为中心向四周形成簇状分支, 每个分支继续进行多级的树状分叉, 分支末端细短, 呈穗状。肠道是整个消化道中长度占比最大的器官, 而小肠又是肠道中长度最长的; 靠近幽门位置的小肠管径最大, 随后逐渐变细。直肠较短, 长度约为小肠的1/7, 管径大, 与小肠前肠部分相近。整个肠道以肠系膜相互连接, 呈盘叠状, 外表面覆盖有淡黄色透明脂肪层。
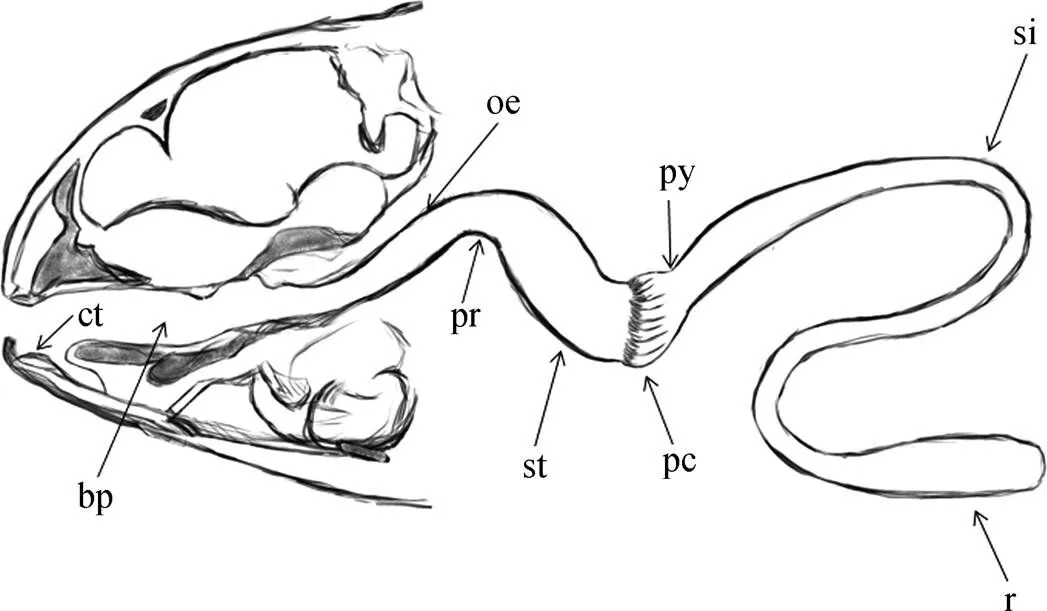
图1 斑石鲷消化道示意图
ct. 臼齿; bp. 口咽腔; oe. 食道; pr. 贲门; st. 胃; py. 幽门; pc. 幽门盲囊; si. 小肠; r. 直肠
ct. cheek tooth; bp. cavum oropharyngeum; oe. oesophagus; pr. preventriculus; st. stomach; py. pylorus; pc. pyloric caecum; si. small intestine; r. rectum
2.2 斑石鲷消化系统组织学特征
2.2.1 食道
斑石鲷食道连接口咽腔和胃, 长度短, 管径大。由外表面向内, 食道依次可分为浆膜层、肌肉层、黏膜下层和黏膜层, 共计4层组织。浆膜层覆盖于食道最外层, 由外周间皮和结缔组织构成。肌肉层位于浆膜层下, 分为明显的两层, 脂肪、血管和结缔组织穿插其中, 外层为环肌, 厚度大, 内层为纵肌, 较薄, 其间夹杂有斜肌。肌肉层下方为黏膜下层, 是一层结缔组织, 内部血管丰富。最内层为黏膜层, 其表面密布突起, 形成数量众多的纵褶, 杯装细胞密集分布于褶皱的表层(图2a、2b)。

图2 斑石鲷消化系统组织结构
a. 食道(10x); b.食道(40x); c.胃(10x); d.胃(40x); e.小肠(10x); f.小肠(40x); g.直肠(10x); h.直肠(40x); i. 肝(40x); j. 胰脏(40x); k.胰脏(40x); oe. 食道; cm. 环肌; lm. 纵肌; ct. 结缔组织; gc. 杯状细胞; se. 浆膜; g. 胃腺; iv. 小肠绒毛; m. 肌肉; rv. 直肠绒毛; g. 胃腺; si. 血窦; va. 空泡; Hd. 肝管; pi. 胰岛; bc. 血细胞; zg. 酶原物质
a. esophagus(10x); b. esophagus(40x); c. stomach(10x); d. stomach(40x); e. small intestine(10x); f. small intestine(40x); g. rectum(10x); h. rectum (40x); i. liver(40x); j. pancreas(40x); k. pancreas(40x);oe. esophagus; cm. circular muscle; lm. longitudinal muscle; ct. connective tissue; gc. goblet cell; se. serosa; g. gastric gland; iv. intestinal villi; m. muscle; rv. rectal villus; si. sinus; va. vacuole; Hd. hepatic duct; pi. pancreas islet; bc. blood corpuscle; zg. zymogen
2.2.2 胃
斑石鲷的胃分为贲门部、胃体部、幽门部3部分, 其中胃体部最为膨大, 贲门部次之。胃组织存在4层结构, 由内及外分别为: 黏膜层、黏膜下层、肌肉层和浆膜层。黏膜层褶皱丰富, 是主要的功能组织; 其中胃体部的褶皱密度最大, 贲门部次之, 幽门部密度最低; 黏膜褶皱表层拥有明显的纹状缘, 褶皱黏膜层中分布有大量胃腺和胃小凹以及少量的杯状细胞。胃腺位于黏膜层下部, 呈管柱状, 相互之间大小存在明显差异。黏膜下层内主要为成分复杂的结缔组织, 密集的血管和淋巴组织在其中穿插, 脂肪组织填充于间隙内。胃的肌肉层极为发达, 尤以幽门部肌肉层为最; 与其他部位的肌肉层相似, 胃的肌肉层也分为环肌和纵肌两层; 内层为环肌, 厚度大, 外层为纵肌厚度相对小, 两者之间以斜肌过渡, 其中夹杂些许脂肪、结缔、血管和神经组织。浆膜膜层由外周间皮包裹少量结缔组织构成, 是胃的最外层(图2c、2d)。
2.2.3 小肠
斑石鲷小肠部分大致可分为前肠、中肠、后肠3部分, 组织结构相同, 分别是黏膜层、黏膜下层、肌肉层和浆膜层。黏膜层内分布有密集的肠绒毛, 肠绒毛上分布有数量众多的杯装细胞, 绒毛外缘内存在大量嗜红颗粒。黏膜下层主要由结缔组织构成, 内部富含血管, 血细胞充盈。小肠的肌肉层相对胃较薄, 且前中后肠之间存在着较为明显的差异; 前肠拥有最厚的肌肉层, 中肠比后肠略厚; 肠的肌肉层结构相同, 最内层为较薄的内纵肌, 中间为较厚的中环肌, 最外层为次厚的外纵肌; 肌层间神经、血管和结缔组织零星可见。肠道最外层浆膜层与胃部类似, 由外周间皮包裹结缔组织形成(图2e、2f)。
2.2.4 直肠
与小肠相同, 直肠也由黏膜层、黏膜下层、肌肉层和浆膜层构成。相比小肠, 直肠的黏膜层肠绒毛更加稀疏, 绒毛长度也较短; 直肠的肌肉层厚度与小肠的前肠部分相似, 较为发达; 黏膜下层的结缔组织相比小肠也更为稀疏; 最外层的浆膜层则依旧为包裹结缔组织的外周间皮(图2g、2h)。
2.2.5 消化腺
斑石鲷的肝脏由大量的肝细胞聚集而成, 细胞呈近似的圆形, 细胞核接近正圆且染色较深; 肝细胞彼此连接形成粗细不均且带有不规则分叉的类网状结构, 细胞间隙内存在血细胞, 其中较大的间隙形成血窦, 内部充满血细胞, 与内部空旷的肝管、胆管差异明显。数量众多的肝细胞中存在透明的空泡结构(图2i)。
斑石鲷的胰脏以弥散状态分布于胃肠之间的缝隙, 从功能和外形上可大致分为两个部分: 内分泌部和外分泌部。外分泌部的胰细胞多数呈柱状或类梭状, 彼此之间界线模糊, 多数胰细胞靠近细胞核的一侧聚集有大量的嗜红颗粒。内分泌部的细胞聚集度高, 呈一个明显的团状, 细胞内存在高浓度的嗜红颗粒, 整体呈现比外分泌部更深的红色(图2j、2k)。
2.3 肠道绒毛
斑石鲷肠道内存在数量众多的指状绒毛, 不同位置的肠绒毛从数量、高度和宽等方面都存在着差异。肠绒毛的高度方面, 小肠前肠肠绒毛的高度明显高于小肠的其他部分和直肠, 小肠中肠部分仅次于前肠, 后肠和直肠最小, 且两者之间的差异不显著。肠绒毛宽度方面, 依然是小肠的前肠部分占据优势显著大于后肠和直肠, 中肠小于前肠但不显著, 后肠和直肠的宽度最窄且与中肠的差异不显著。肠绒毛数量方面, 小肠前肠数量最多, 且与其他部分有着显著差异; 中肠和剩余部分的差距与宽度数据类似, 有差距但不如高度数据显著; 后肠和直肠依旧最低, 且两者之间的差异不显著(图3)。
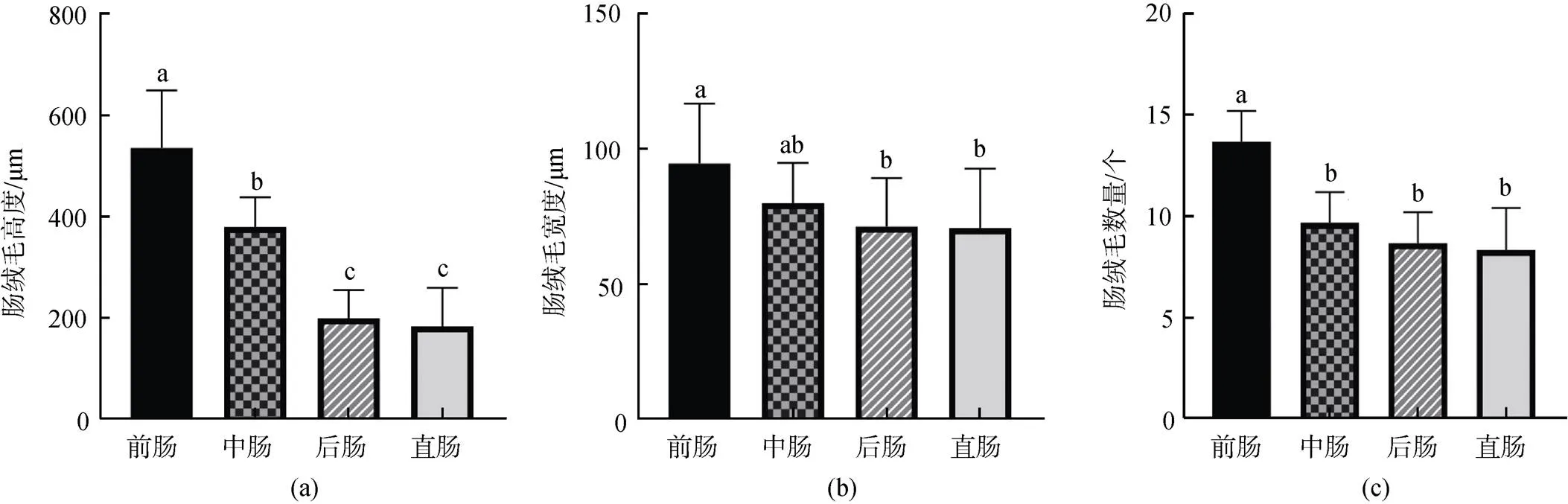
图3 斑石鲷肠道绒毛高度、宽度、数量对比
不同上标间存在显著差异,<0.05
Significant differences indicated by different superscripts,< 0.05
3 讨论
3.1 消化道形态特征与食性的关联
斑石鲷的野生种群主要分布于中国沿海, 日本和韩国的部分海域亦有分布; 属于近海温水鱼类, 主要以礁岩间栖息的小型贝类和甲壳动物为食, 在食性上偏向肉食性。鱼类消化道的形态学特点往往表现出与种属、食性以及生活环境的息息相关[10]。斑石鲷消化道的形态学特征与食性特点的对应很好的体现了这一点。斑石鲷上下颌齿整体愈合, 呈鹦鹉喙状, 整体前凸, 适合在礁石间觅食海胆、贝壳等软体动物; 口咽腔内部宽大的空间, 还有颌齿后侧两排坚硬的臼齿, 能够对进入口咽腔的食物进行粉碎、研磨, 从而降低食物的颗粒度。鳃耙上分布的棘状齿能够对粉碎、研磨后的食物进行筛选, 滤出其中如贝壳、甲壳等坚硬且难以消化的食物碎片并吐出, 尽可能地避免杂质损伤后续的消化道。口咽腔的种种特征都表现出斑石鲷为摄食贝类、甲壳类及软体动物所做出的特异性进化。食道长度短, 管径粗, 内部密布的纵褶说明其拥有极强的扩张和收缩的能力, 这使得斑石鲷能够吞下大块的食物团, 并对其进行一定的塑形, 从而更加顺畅的进入胃部[11]。V型的胃是典型的肉食性鱼类特征, 内部的容积大, 能够让更多的食物驻留更长的时间, 使斑石鲷拥有较长的摄食间隔。幽门盲囊分支多且密集, 能够分泌多种消化酶, 有效提高斑石鲷的营养吸收效率[12]。肠道系数作为衡量鱼类形态的重要参数, 其数值的大小往往和鱼类的食性息息相关[13]。本次实验中的斑石鲷平均肠道系数为1.94, 大于牙鲆(0.57) ()、蓝点马鲛鱼(0.56)()[14]等典型的肉食鱼类, 小于鳙(4.63~4.97)()[15]、湘华鲮(21.42)()[16]、黄尾密鲴(6.25)()[17]等典型的植食性鱼类。从斑石鲷明显的胃组织以及明显长于体长的肠道, 可以比较容易地推断出斑石鲷是一种杂食偏肉食性的鱼类。
3.2 消化系统组织学特征与功能的关联
斑石鲷食道短粗, 由内至外可分为4层: 黏膜层、黏膜下层、肌肉层和浆膜层。黏膜层表面存在数量众多的纵褶, 数量众多的杯状细胞密集的分布于黏膜层中。在吞咽食物时通过对纵褶的舒展来调整食道管径的大小, 杯状细胞分泌的黏液覆盖包裹在食物表面, 避免大体积的食物和其中的坚硬物质对黏膜造成损伤。杯状细胞除了润滑食物外[18], 还具有一定的吸收和转运大分子、辅酶因子, 抵御外来致病因子等作用[19]。斑石鲷的食物中甲壳动物和软体动物占了多数, 在吞咽这些食物时难免夹杂这些生物的坚硬外壳, 杯状细胞所分泌的黏液不仅能有效缓解斑石鲷食道损伤产生的可能, 对于已经产生的损伤也能起到很好的保护作用。食道肌肉层大致可分为两层, 靠近内侧厚实的环肌, 和靠近外侧的稍薄的纵肌, 两种肌肉彼此配合, 将食物输送至胃腔。黏膜下层由结缔组织构成, 其间分布有密集的血管和神经组织, 为食道的肌肉活动提供支持。作为以有壳动物为主食的生物, 斑石鲷无胃内的消化液有着相比其他动物更强的消化能力, 而厚实的肌肉层确保了斑石鲷食道的封闭性, 能有效阻止胃部的消化液的反流, 在确保胃腔内消化液浓度的同时也能保护食道和口咽腔的黏膜免受消化液的侵蚀。
胃部作为整个消化道最为膨大的器官, 与食道相似, 整体结构也分为4层: 黏膜层、黏膜下层、肌肉层和浆膜层。黏膜层褶皱丰富, 胃腺密集, 胃基底部这种现象尤为明显。胃部的黏膜下层较之其他消化道也更加厚, 密集交错的神经和血管网络穿插中, 为胃腺和胃肌的相关活动提供着足够的物质支持[20]。胃拥有整个消化道最为发达的肌肉层, 而肌肉层的厚度往往与食物的机械消化能力息息相关[21]。丰富的黏膜褶皱极大增加了与食物的接触面积, 密集的胃腺分泌出足够的消化液, 厚实发达的肌肉将消化液和食物充分的搅拌混合, 而较大的胃腔体积和及其发达的幽门括约肌让食物能在其中停留较长的时间, 从而使得食物中的动物蛋白能够被充分地消化、分解。斑石鲷多以贝类、虾蟹等动物性饵料为食物来源, 与胃的组织学特征相对应。斑石鲷胃部的组织学特征与马苏大马哈鱼这样的典型的肉食性鱼类相似, 胃组织明显且发达, 与典型的植食性鱼类存在明显差异, 如长麦穗鱼并不存在明显的胃组织[5, 22]。
肠道也拥有着和胃、食道相似的结构, 不同的是肠道的黏膜下层极薄, 几乎与黏膜层融合。肠道黏膜层上分布有数量众多的肠绒毛, 其中前肠处的肠绒毛密度最高, 绒毛长度最长; 随后便向着后肠方向逐级降低, 后肠肠绒毛的长度和密度都远小于前肠。肠绒毛的数量和长度与鱼类的消化吸收能力息息相关[23, 24], 由此可以推断, 营养的消化吸收主要在斑石鲷的前、中肠进行。此外, 整个肠道黏膜层中还分布有数量众多的杯装细胞, 其分泌的黏液不仅对食糜的流动起到润滑的作用, 同时也对黏膜层形成了保护, 并对帮助转运食糜中的大分子物质[13]。黏膜下层虽然厚度薄, 但是其内部的血管密度远高于胃和食道, 大量的血细胞几乎充斥整个黏膜下层, 在前肠中体现的最为明显。密布的血管和丰富的血细胞能够将肠黏膜吸收的营养物质以最高的效率转运至鱼体全身, 进一步证明了肠道在物质消化吸收中的主体地位。斑石鲷的肠道相对较短, 而所摄食的食物相比植食性鱼类营养密度更高, 因此需要在较短的时间内吸收更多的营养物质, 黏膜下层高密度的血管及丰富的血细胞正是为了满足这一需求的表现。肠道肌肉层的厚度依然遵循从前肠至后肠递减的规律。前肠较厚的肌肉层在帮助食糜流动的同时, 也对食糜施加着更大的压力, 从而让其与肠黏膜进行更充分的接触, 增加营养吸收的效率。直肠黏膜层中绒毛稀疏且高度较低, 仅承担水分等营养物质的重吸收[13], 数量丰富的杯装细胞和厚实的肌肉层让直肠的蠕动更加顺畅, 有利于食物残渣的排泄。此外由于更接近外部环境, 充足的黏液细胞也能起到屏障的作用, 避免病原体的侵袭。斑石鲷食物的特殊性就决定了其消化完毕后的残渣中依然会存在哪些有壳动物的坚硬组织, 在更强的肌肉压力下, 需要足量杯装细胞分泌的黏液下才能减少直肠黏膜的损伤。
斑石鲷的肝脏中出现的空泡结构均匀分布在绝大多数的肝细胞中, 且大小相似, 其中的物质是肝糖原, 主要用于帮助斑石鲷调节血糖水平和抵御短暂饥饿[25]。肝血窦连接着门静脉和肝动脉, 将血液中的氧气和营养物质输送至肝脏中, 同时将肝脏分泌物运送到鱼体各处[26]。胰脏细胞内深色的嗜红颗粒为酶原物质, 其中多为各种消化酶的前体, 在进入肠道后被相应的环境因子激活, 形成相关的催化能力[27]。胰脏中呈团状的胰岛组织属于内分泌部, 主要分泌各种激素, 功能大多与调节血糖水平相关[28], 部分也有调节机体生长的作用[29]。
综上, 斑石鲷消化系统的各组织学特征与各组织器官的承担的消化功能相对应。其具有与肉食性鱼类相似的胃组织的同时肠道长度更长, 但又明显短于典型的植食性鱼类,, 综合特征判断其属于杂食偏肉食性。斑石鲷口咽腔中用于碾碎甲壳的臼齿, 食道和直肠的肌肉发达, 黏膜中杯装细胞丰富, 这些都是斑石鲷为摄食甲壳和软体动物做出的适应性改变。
[1] 孟庆文, 苏锦祥. 鱼类分类学[M]. 北京: 中国农业出版社, 1995: 734-756.
MENG Qingwen, SU Jinxiang. Taxonomy of fishes[M]. Beijing: China Agriculture Press, 1995: 734-756.
[2] 高小强, 洪磊, 黄滨, 等. 斑石鲷种苗繁育与养殖技术研究进展[J]. 水产研究, 2018, 5(2): 112-122.
GAO Xiaoqiang, HONG Lei, HUANG Bin, et al. Research progress in seedling breeding and breeding technology of[J]. Fisheries Research, 2018, 5(2): 112-122.
[3] 楼允东. 组织胚胎学(第二版)[M]. 北京: 中国农业出版社, 1998: 95-114.
LOU Yundong. Histoembryology (2nd Ed)[M]. Beijing: China Agriculture Press, 1998: 95-114.
[4] 尤宏争. 鱼类消化系统及消化酶的研究进展[J]. 河北渔业, 2012, 3: 95-114.
YOU Hongzheng. Research progress of fish digestive system and digestive enzymes[J]. Hebei Fisheries, 2012, 3: 95-114.
[5] 杨晓鸽, 连玉喜, 万安, 等. 长麦穗鱼消化系统的形态学与组织学研究[J]. 水产学杂志, 2021, 34(2): 45-50.
YANG Xiaoge, LIAN Yuxi, WAN An, et al. Morphological and histological studies on the digestive system of[J]. Chinese Journal of Fisheries, 2021, 34(2): 45-50.
[6] 胡玲玲, 李加儿, 区又君, 等. 养殖条石鲷消化道形态构造及组织学观察[J]. 南方水产, 2010, 6(6): 65-69.
HU Lingling, LI Jiaer, OU Youjun, et al. Study onmorpholigy and histology of digestive tract in cultured[J]. Southern Fisheries, 2010, 6(6): 65-69.
[7] 曹潇, 孙敏, 朱亮华, 等. 黑鱾消化系统的形态学及组织学研究[J]. 热带海洋学报, 2018, 37(1): 20-26.
CAO Xiao, SUN Min, ZHU Lianghua, et al. Morphology and histology of the digestive system in, 2018, 37(1): 20-26.
[8] 王雨福. 斑石鲷()早期生长特征及消化系统发育的研究[D]. 青岛: 中国科学院海洋研究所, 2015.
WANG Yufu. Early growth characteristics and development of digestive system of[D]. Qingdao: Institute of Oceanology, Chinese Academy of Sciences, 2015.
[9] 陈国华, 王永波, 王珺, 等. 波纹唇鱼消化系统的组织学[J]. 水生生物学报, 2010, 34(4): 685-693.
CHEN Guohua, WANG Yongbo, WANG Jun, et al. Histology of the digestive system in[J]. Acta Hydrobiologica Sinica, 2010, 34(4): 685-693.
[10] 赵彦花, 区又君, 李加儿, 等. 黄唇鱼消化系统组织结构及黏液细胞分布特征[J]. 渔业科学进展, 2019, 40(3): 80-86.
ZHAO Yanhua, OU Youjun, LI Jiaer, et al. Histology and distribution of mucous cells in digestive system of[J]. Advances in Fisheries Science, 2019, 40(3): 80-86.
[11] NEBEL O T, CASTELL D O. Lower esophageal sphincter pressure changes after food ingestion[J]. Gastroenterology, 1972, 63(5): 778-783.
[12] PHAM V H, BENJAKUL S. Purification and characterization of trypsin from pyloric caeca of bigeyesnapper ()[J]. Journal of Food Biochemistry, 2006, 30(4): 478-495.
[13] 殷名称. 鱼类生态学[M]. 北京: 中国农业出版社, 1995: 64-78.
YIN Mingcheng. Fish Ecology[M]. Beijing: China Agriculture Press, 1995: 64-78.
[14] 王迎春, 周勤, 段晓英. 八种海产硬骨鱼类消化系统的比较解剖研究[J]. 海洋湖沼通报, 1997, 3: 46-51.
WANG Yingchun, ZHOU Qin, DUAN Xiaoying. Comparative anatomical study on digestive system of eight Marine teleost fishes[J]. Bulletin of Oceanology and Limnology, 1997, 3: 46-51.
[15] 林浩然. 五种不同食性鲤科鱼的消化道[J]. 中山大学学报(自然科学版), 1962, 3: 65-78, 93-95.
LIN Haoran. Digestive tract of five different carnivorous cyprinidae[J]. Journal of Sun Yat-sen University (Natural Science), 1962, 3: 65-78, 93-95.
[16] 梁志强, 李传武, 刘明求, 等. 湘华鲮消化系统的形态学与组织学研究[J]. 中国水产科学, 2011, 18(5): 1051-1060.
LIANG Zhiqiang, LI Chuanwu, LIU Mingqiu, et al. Morphological and histological observation of the digestive system in(Nichols)[J]. Journal of Fishery Sciences of China, 2011, 18(5): 1051-1060.
[17] 徐如卫, 吴振兴, 罗仙池. 新安江水库黄尾密鲴Bleker生物学研究[J]. 浙江水产学院学报, 1996, 3: 20-32.
XU Ruwei, WU Zhenxing, LUO Xianchi. Studies on the biology characteristics ofBleker in Xin’anjiang Reservoir[J]. Journal of Zhejiang Fisheries University, 1996, 3: 20-32.
[18] 马爱军, 马英杰, 姚善诚. 黑鲷消化系统的胚后发育研究[J]. 海洋与湖沼, 2000, 31(3): 281-287.
MA Aijun, MA Yingjie, YAO Shancheng. Studies on post-embryo development of the digestive system of[J]. Oceanologia et Limnologia Sinica, 2000, 31(3): 281-287.
[19] ARELLANO J M, STORCH V, SARASQUETE C. Ultrastructural study on the intestine of Senegal sole,[J]. Journal of Applied Ichthyology, 2010, 18(3): 154-158.
[20] 沈雷, 孙石柱, 刘冰华, 等. 胃黏膜下层的动脉构筑及其临床意义[J]. 齐齐哈尔医学院学报, 2008, 19: 2311- 2322.
SHEN Lei, SUN Shizhu, LIU Binghua, et al. Arterial architecture of gastric submucosa and its clinical significance[J]. Journal of Qiqihar Medical College, 2008, 19: 2311-2322.
[21] Trevor A. Histological and cytological structure of the gastrointestinal tract of the luderick,(pisces, kyphosidae), in relation to diet[J]. Journal of Morphology, 1986, 190(1): 109-119.
[22] 李培伦, 王继隆, 唐富江. 马苏大麻哈鱼幼鱼消化系统形态学和组织学观察[J]. 水产学杂志, 2021, 34(4): 43-48.
LI Peilun, WANG Jilong, TANG Fujiang, et al. Morphological and histological observation of digestive system of juvenile Masu Salmon[J]. Chinese Journal of Fisheries, 2021, 34(4): 43-48.
[23] 威尔士·斯托赫. 比较动物细胞学和组织学[M]. 北京: 科学出版社, 1979: 217-245.
Wales Stoch. Comparative animal cytology and histology[M]. Beijing: Science Press, 1979: 217-245.
HUANG Zihao, YI Jianhua, CHEN Dongming, et al. Morphological and histological observations of digestive system in[J]. Freshwater Fisheries, 2015, 45(2): 8-14.
[25] BOULHIC M, GABAUDAN J. Histological study of the organogenesis of the digestive system and swim bladder of the Dover sole,(Linnaeus 1758)[J]. Elsevier, 1992, 102(4): 373-396.
[26] BRUNT E M, GOUW A S H, HUBSCHER S G, et al. Pathology of the liver sinusoids[J]. Histopathology, 2014, 64(7): 907-920.
[27] MILLER S A J, BRUCE L. Harrison’s principles of internal medicine, 19th edition[J]. Confusion and Delirium, 2015, 2437-2438.
[28] KELLY C, NEVILLE H, et al. Role of islet structure and cellular interactions in the control of insulin secretion[J]. Islets, 2011, 3(2): 41-47.
[29] AMISTEN S, SALEHI A, RORSMAN P, et al. An atlas and functional analysis of G-protein coupled receptors in human islets of Langerhans[J]. Pharmacology & Therapeutics, 2013, 139(3): 359-391.
Morphological and histological observations of thedigestive tract
CHEN Xiao1, 2, 3, 4, XIAO Zhi-zhong1, 8, 9, XIAO Yong-shuang1, 4, JI Guang-lei5, DENG Qin-you2, 3, LI Xuan2, 3, WANG Yu-fu1, 4, MA Yu-ting1, 4, 5, 6, LI Jun1, 4, LIU Xiang-quan2, LIAN Chang7, LIAN Ya-ming8, YANG Li-zhong9
(1. CAS/Shandong Key Laboratory of Experimental Marine Biology, Institute of Oceanology/Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China; 2. Shandong Provincial Key Laboratory of Restoration for Marine Ecology, Shandong Marine Resource and Environment Research Institute, Yantai 264006, China; 3. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China; 4. Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao Laboratory for Marine Biology and Biotechnology, Qingdao 266237, China; 5. Institute of Ocean Development of Weihai, Weihai 264200, China; 6. College of Marine Science and Engineering, Qingdao Agricultural University, Qingdao 266109, China; 7. Weihai Wendeng Haihe Aquatic Nursery Co., Weihai 264400, China; 8. Weihai Haohuigan Marine Biotechnology Co., Weihai 264400, China; 9. Putian Pusheng Aquatic Science and Technology Co., Putian 351174, China)
The objectives of this study were to examine the morphological and histological characteristics of thedigestive system and provide basic data for the artificial culture of. We conducted relevant studies on thedigestive system using anatomy and paraffin sections. The digestive tract was composed of six parts, including the oropharyngeal cavity, the esophagus, the stomach, the pyloric cecum, the small intestine, and the rectum.had a large space inside the oropharyngeal cavity, and its jaw teeth were connected and fused, similar to a parrot’s beak. Pharyngeal teeth are like flat round shields distributed inside the jaw teeth. This structure allowsto crush a crustacean shell and eat the meat inside. The esophagus is short and thick, and there are many longitudinal folds on the mucosal layer and several goblet cells in the mucosa. These folds helpswallow food smoothly. The stomach is asymmetrical and V-shaped with dense gastric glands under the mucosal layer, and the pylorus has the thickest muscle layer in the entire digestive tract. This is where most of the foodswallows is digested and metabolized. The small intestine is divided into three parts: the foregut, the midgut, and the hindgut. Their pipe diameter decreases in turn. The density and length of the intestinal villi in the mucosal layer follow the same rule. The nutrient absorption function ofis mainly concentrated in the foregut and midgut based on the intestinal structure. The rectum is similar in diameter to the anterior segment of the small intestine but is only one-seventh of the length. The intestinal villi in the rectum are shorter in length and less dense than those in the small intestine. The rectum is responsible for reabsorbing water and trace elements from the food residue, and the residue is compressed and excreted. The apparent simplicity of its structure corresponds to a simple function.
; digestive tract; morphology; histologic
Jan. 6, 2022
[National Key R&d Program of China, No. 2018YFD0901204; Putian City Science and Technology Plan-STS Project Supporting the Cooperation between the University and the City, No. 2019HJSTS010; Shandong Agricultural Improved Seed Project, No. 2017LZGC009]
S961
A
1000-3096(2022)07-0070-08
10.11759/hykx20220106003
2022-01-06;
2022-04-22
国家重点研发计划项目(2018YFD0901204); 莆田市科技计划-STS计划配套院市合作项目(2019HJSTS010); 山东省农业良种工程(2017LZGC009)
陈宵(1994—), 男, 江苏扬州人, 硕士研究生, 主要从事鱼类繁育研究, 电话: 18013973189, E-mail: 1027454514@qq.com; 刘相全(1968—),通信作者, 电话: 18153518269, E-mail: lxq6808@163.com; 李军(1964—), 通信作者, 电话: 0532-82898718, E-mail: junli@qdio.ac.cn
(本文编辑: 谭雪静)
