内耳支持细胞促进羊水干细胞定向分化为神经元的可能调控机制研究
2022-06-15宗凌姜鸿彦
宗凌姜鸿彦
1广州医科大学附属第二医院耳鼻咽喉科(广州 510260)
2海南省人民医院耳鼻咽喉医院(海口 570311)
内耳螺旋神经元损伤导致的神经性耳聋,目前缺乏有效的预防、治疗措施,而干细胞移植可能成为治疗神经系统损伤的一个有效手段[1-4]。前期研究证实:羊水干细胞(amniotic fluid stem cells,AFS)具有向神经元定向分化的潜能,未经诱导(92.0±1.0%)表达神经元标记物Tuj1(anti-Neuronal Class III β-Tubulin(Tuj1)antibody),但缺乏神经元形态特征[5];而将人来源的AFS分别接种于小鼠基底膜和前庭组织来源的饲养层形成支持接触-立体三维共培养模式,即使不添加神经生长因子和信号诱导因子,仅用DMEM/F12作为培养基,约37-52%的AFS可定向分化为听觉神经元[6,7];AFS经悬滴培养形成拟胚体,具有神经干性,与耳蜗基底膜共培养(二维培养模式,无立体支持接触),并不能诱导拟胚体向形态典型的神经元分化[8],说明支持接触-立体三维共培养模式是AFS分化为听觉神经元的前提条件,而了解其发生和调控机制,对于螺旋神经元再生,及AFS豚鼠耳蜗内移植的替代研究,及后续的临床应用具有重要的现实意义。
立体三维接触是AFS及其他干细胞有效分化为神经元/毛细胞的前提条件[9-11],说明这种模式参与干细胞定向分化及调控,而内耳支持细胞对AFS的分化作用仍不明确。文献报道结合我们前期研究推测AFS本身就具有特殊性,不仅表达间充质干细胞标记物,也表达神经干/祖细胞标记物,并具有向神经元自发分化的趋势,提示AFS在向神经元方向诱导上可能较其他种子细胞有优势[5-7]。另外,分别获取前庭和基底膜来源的内耳支持细胞,发现前庭来源的饲养层分化效果要优于基底膜[6,7],说明二者的内在差异可能也参与神经元分化的调控。在哺乳类Corti器支持细胞群中存在P27kip1,其在于促使细胞退出细胞周期,停止分裂,保持不能增殖的状态[12];前庭和基底膜来源的饲养层细胞都表达P27kip1[5,6],而两者诱导神经分化能力的差异是否与P27kip1表达量相关,仍有待研究。
实验拟从AFS接种密度,内耳支持细胞P27kip1表达差异入手,研究上述因素与AFS向神经元定向分化的潜在联系。
1 材料和方法
1.1 实验材料
1.1.1 实验小鼠:出生1~3d的C57BL/6J小鼠30-50只(雌雄不限),由中山医学院实验动物中心提供。
1.1.2 羊水标本:由中山大学附属第一医院提供,孕19~22周,经B超引导下抽取羊水10-15ml。羊水取自产前检查家庭,夫妻双方均签署知情同意书,研究经中山大学附属第一医院医学伦理委员会审核批准。
1.1.3 主要试剂:DAPI一抗、Tuj1一抗、P27kip1一抗购于美国Abcam公司。
1.2 实验方法
1.2.1 分离并培养扩增AFS
将羊水接种于25cm2的培养瓶,第7天或第8天,根据羊水贴壁细胞的增殖情况,全量换液,继续培养2~3d,观察羊水贴壁细胞的生长状态,决定是否需要传代[4-7]。采用德国Miltenyi Biotec公司的c-Kit磁珠抗体,分选羊水中具有c-Kit标记的AFS[6,13]。
1.2.2 基底膜和前庭来源的内耳支持细胞饲养层的制备
解剖显微镜下,分离基底膜、椭圆囊和球囊组织,经酶解消化细胞培养6天;收集悬浮的内耳干细胞空心球体,再次酶解消化,用DMEM/F12重悬细胞,按1×105/cm2的密度接种于无菌盖玻片上,免疫荧光鉴定P27kip1的表达[6,7]。
1.2.3 内耳支持细胞对AFS(不同密度接种)的定向分化
制备基底膜来源的内耳支持细胞饲养层,24小时后,将细胞培养液弃去,将AFS消化、离心,用DMEM/F12 重悬细胞,分别将 2×105、4×105、6×105、8×105、10×105个细胞接种在制备好的饲养层上培养6天,检测Tuj1的表达[6];
1.2.4 内耳支持细胞(基底膜和前庭来源)对AFS的定向分化
分别选取基底膜和前庭组织来源的空心球体,按接种密度1×105/cm2制备内耳支持细胞饲养层,免疫荧光检测这两种饲养层细胞P27kip1的表达;将10×105个AFS分别接种于这两种饲养层上,用DMEM/F12培养6天,检测Tuj1的表达[6,7]。
1.2.5 细胞计数
免疫荧光染色 Tuj1、P27kip1、DAPI,以细胞突起长度为胞体直径5倍以上的神经样细胞表型作为形态学评价指标,使用Leica显微镜观察;分别选取5个样本,每个样本随机选取4个不同视野,计数20倍镜视野下观察到的Tuj1、P27kip1阳性细胞数与DAPI阳性细胞数之比,计算20个视野的平均值。
1.3 统计学处理
阳性细胞表达率以均值±标准差表示,采用SSPSS 17.0软件进行统计学分析,组间比较采用Oneway ANOVA检验,P<0.05为差异具有统计学意义。
2 结果
2.1 AFS接种密度相关性
分别将2×105、4×105、6×105、8×105、10×105个AFS接种在基底膜来源的饲养层表面,经支持接触-立体三维共培养6天,AFS可以分化为Tuj1阳性且形态典型的神经元。AFS以高密度(10×105)接种于饲养层能产生更多形态典型的神经元(20倍镜视野下,平均208±25.7条神经突起),相对于低密度(2×105)接种(20倍镜视野下,平均41±12.3条神经突起)(图1);随着接种细胞数的增加,神经突起的数量增多(P<0.01,n=10),神经突起的长度基本保持不变,平均88±7.8μm(P>0.05,n=10)(图2)。
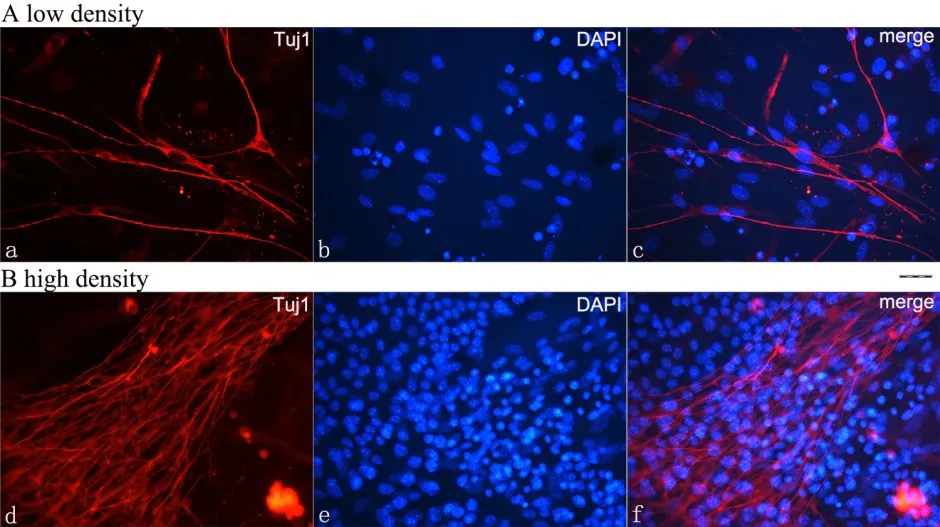
图1 AFS分别以高密度(10×105)和低密度(2×105)接种于饲养层细胞表面。A为低密度接种;B为高密度接种;a,d红色荧光代表Tuj1;b,e 蓝色荧光代表DAPI;c,f叠加图像,标尺=25μm。Fig.1 The AFS were inoculated on the feeder layer with low density(2×105)(A)and high density(10×105)(B),respectively.a,d red fluorescence represents Tuj1;b,e Blue fluorescence represents DAPI;c,f Overlay image;Bar=25μm.
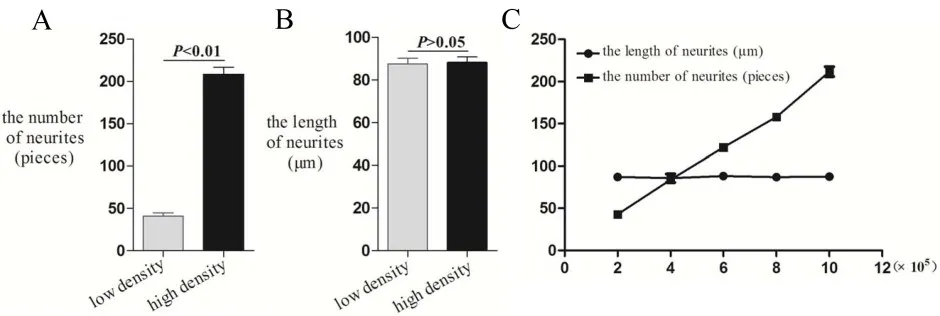
图2 AFS分别以2×105、4×105、6×105、8×105、10×105接种于饲养层表面,低密度(2×105)和高密度(10×105)相比较,高密度分化所得的神经突起数量增多(P<0.01,n=10,A),神经突起长度无统计学差异(P>0.05,n=10,B);C神经突起数量随着接种比例的增加而逐渐上升;神经突起长度随着接种比例的增加,基本保持不变。Fig.2 The AFS were inoculated on the feeder layer with 2×105,4×105,6×105,8×105and 10×105,respectively.2×105represents low density.10×105represents high density.The number of neurites was significantly higher on high density than low density(P<0.01,n=10,A),the length of neurites have no statistical differences(P>0.05,n=10,B).With the increase of inoculation proportion,the number of neurites gradually increased,but the length of neurites remained unchanged(C).
2.2 内耳支持细胞P27kip1表达差异对神经元分化的影响
将基底膜和前庭组织来源的空心球体制备饲养层,都表达内耳支持细胞标记物P27kip1;前庭P27kip1(79.8±8.0%),明显高于基底膜(46.9±9.7%)(P<0.01,n=10,图3)。

图3 基底膜与前庭来源的饲养层细胞都表达P27kip1。A基底膜,B 前庭,a.d P27kip1,b.e DAPI,c.f叠加图像;C 基底膜与前庭P27kip1表达量对比,前庭P27kip1明显高于基底膜(P<0.01,n=10),标尺=50μm。Fig.3 The P27kip1was on the feeder layer cells derived from the Corti’s organ(A)and vestibular tissue(B).a,d P27kip1;b,e DAPI;c,f Overlay image;(C)the comparison of P27kip1between the Corti’s organ and vestibule tissue.The P27kip1on the vestibule tissue was significantly higher than the Corti’s organ(P<0.01,n=10);Bar=50μm.
而将AFS种植于前庭组织来源的内耳支持细胞饲养层,分化得到的神经突起数量(20倍镜视野下,平均81.1±15.6条)和神经突起长度(20倍镜视野下,平均110.9±11.9μm),都优于基底膜(20倍镜视野下,平均41.4±12.9条,平均长度90.1±12.9μm,P<0.01,n=10,图4);

图4 AFS分别接种于基底膜(A)和前庭(B)组织来源的内耳支持细胞饲养层。a.d Tuj1,b.e DAPI,c.f叠加图像;与基底膜相比,前庭来源的饲养层分化所得的神经突起数量和长度增多(P<0.01,n=10,C,D),标尺=50μm。Fig.4 The AFS were inoculated on the feeder layer derived from the Corti’s organ(A)and vestibular tissue(B).a,d Tuj1;b,e DAPI;c,f Overlay image;The number and length of neurites was significantly higher on vestibular tissue than Corti’s organ(P<0.01,n=10,C,D),Bar=50μm.
基底膜和前庭组织来源的饲养层都表达内耳支持细胞标记物P27kip1,前庭P27kip1表达量明显高于基底膜;前庭组织来源的内耳支持细胞对神经元定向分化的能力,明显强于基底膜组织。统计分析发现P27kip1表达量与分化的神经突起的数量和长度均呈正相关(图5)。
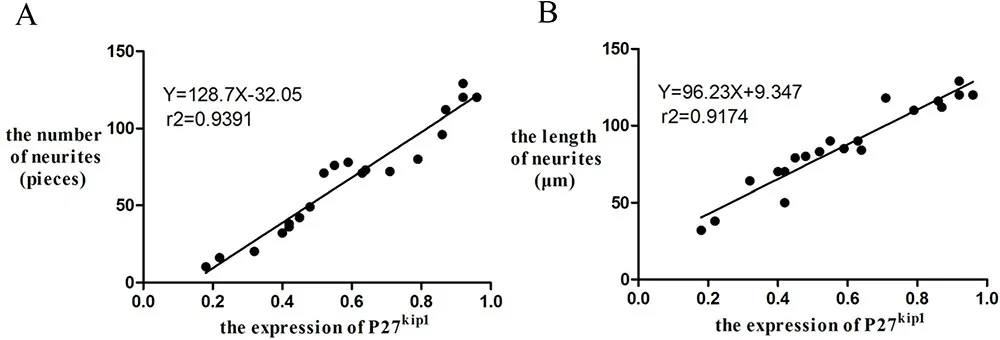
图5 内耳支持细胞饲养层P27kip1表达量与神经突起的数量(A)和长度(B)呈正相关。Fig.5 The P27kip1in the inner ear supporting cells feeder layer was positively correlated with the number(A)and length(B)of neurites.
3 讨论
由螺旋神经元损伤导致的神经性耳聋是耳科常见疾病之一,虽已有文献证实,胚胎干细胞和诱导性多能干细胞在体外能分化为神经元,但由于接种活体动物有致瘤性,限制了此类干细胞的临床应用。AFS具有全能干细胞的特点[13],接种活体动物也无致瘤性。前期研究已证实,AFS有向神经元定向分化的潜能[5],与小鼠基底膜或前庭组织来源的内耳支持细胞共培养,能定向分化为神经元。探寻其定向分化机制,有助于深入神经元分化、再生和保护的相关研究,并对耳科基础研究向临床运用转化具有重要的前期指导意义。
小鼠内耳支持细胞诱导AFS定向分化为神经元的机制迄今并不明确。研究已证实未经诱导的多个不同AFS样本,神经元标记物Tuj1呈强阳性,传代30次后表达仍未减弱,提示AFS在向神经元方向分化上较其他种子细胞可能更具有优势[5]。课题组前期研究已证实内耳支持细胞在体外可能通过Wnt/β-catenin信号通路调控AFS向神经元方向分化[6-8],推测AFS内在Wnt信号参与调控(密度/剂量依赖性)。随着接种比例的提高,更高比例的Wnt信号相关因子参与促进神经元分化。同时,低密度接种AFS已经可以有效地分化为神经元,具有和高密度接种一样的神经突起长度,说明神经突起长度(88±7.8μm)可能已达到稳定/接近成熟状态,或归因于AFS诱导能力的极限。增加AFS数量,可使神经突起数量增加,提高移植效率。
分别获取基底膜和前庭来源的内耳干细胞空心球体制备饲养层,接种的细胞密度都是1×105/cm2,对基底膜和前庭来源的饲养层进行免疫荧光染色,鉴定内耳支持细胞标记物P27kip1,发现虽然接种细胞密度一样,但前庭P27kip1表达量确实比基底膜要高。推测可能前庭制备的饲养层细胞存活率比基底膜要好,因此以相同密度接种,前庭和基底膜P27kip1的表达量差异显著,所以我们才进一步提出假设,可能羊水干细胞向神经元定向分化与P27kip1相关;另外,在小鼠胚胎发育的第13至14天,超过80%的内耳祖细胞精确分化为终末型毛细胞和支持细胞。哺乳动物耳蜗毛细胞损伤后自身不能分裂增生,同时其周围的支持细胞亦保持不能分裂增生的状态,不能转化成毛细胞,而P27kip1在这当中起重要作用[14],与毛细胞和支持细胞祖细胞停止有丝分裂有关;在P27Kip1基因被靶向敲除的小鼠模型中,胚体发育第14天后,感觉祖细胞增殖仍持续存在,导致出现多余的毛细胞和支持细胞,这表明成熟支持细胞中P27Kip1持续表达可能有助于维持该组织的静止状态。另外,Corti器由精确排列的内、外毛细胞,支持细胞等构成;而一排内毛细胞和三排外毛细胞间都有P27Kip1表达的内耳支持细胞相间隔并依托包绕,P27Kip1可能定向诱导与其直接接触的内耳祖细胞退出有丝分裂形成终末型毛细胞。基底膜和前庭组织来源的饲养层都表达内耳支持细胞标志物P27kip1;而将AFS种植在内耳支持细胞表面,模拟耳蜗Corti器的分化过程,发现37-52%的AFS可定向分化为神经元,推测可能是内耳支持细胞P27kip1诱导与其直接接触的AFS停止有丝分裂,退出细胞周期,开始自发的向神经元方向分化[6,7]。前庭来源内耳支持细胞P27kip1的表达量高于基底膜,在与AFS直接接触共培养中,前庭分化得到的神经突起在数量和长度方面均优于基底膜,P27kip1与神经突起的数量和长度呈正相关,推测内耳支持细胞饲养层P27kip1增多,导致更多的AFS停止有丝分裂,失去干性,又由于本身具有向神经元定向分化的特殊性,最终产生大量神经元。
因此,AFS接种密度以及内耳支持细胞P27kip1表达水平与调控AFS神经元定向分化有关。适当增加AFS接种比例、采用前庭来源的内耳支持细胞作为饲养层、有效调控Wnt信号、过表达P27kip1基因等,均有助于AFS向神经元分化,提高效率。这对临床转化研究具有参考价值。
当然,本研究仍存在局限性。深入Wnt信号因子、P27kip1基因的相关研究,分析相应内耳支持细胞饲养层对AFS神经元分化的影响,可能更好阐述其相关机制。
