不同种群巨柏种子表型及萌发特性
2022-05-16李永霞颜呈霖张一鸣杨小林赵垦田
李永霞, 颜呈霖, 张一鸣, 杨小林, 赵垦田
(1.西藏大学理学院, 拉萨 850000; 2.西藏农牧学院资源与环境学院, 西藏 林芝 860000)
种子表型特征是指具有特定基因型的个体,在一定环境条件下所表现出来的性状特征的总和[1]。生物个体所处的特定环境条件与基因同时发挥作用,使得表现型呈多样性。植物种子表型可塑性在一定程度上揭示种源的遗传规律和变异程度,反映着植物的生长环境条件和适应能力[2-3]。种子性状(大小、质量等)作为植物生活史的关键特征,在植物野生种源开发以及选种育种等方面存在较大的研究潜力。
巨柏为柏科(Cupressaceae)柏木属(Cupressus),是西藏特有常绿乔木树种[4],狭窄分布于雅鲁藏布江中游林芝、米林至朗县一带的河谷山地,作为该分布区域的顶级群落,在水土保持、涵养水源和气候调节等方面具有重要生态服务功能,占据重要生态位[5]。由于人为活动及自然因素影响,巨柏个体逐渐减少,天然更新较差,处于濒危状态[6]。种子是巨柏更新的关键环节,基于种子表型特征的优良种质资源筛选是重要的基础性研究。目前,巨柏相关研究主要集中在种群群落特征、遗传多样性、苗木培育和种子萌发[7-13]等方面,而种子表型特征及其变异的研究相对较少。本研究选择不同种群的巨柏种子作为试验材料,分析种子表型特征及萌发特性,为巨柏种质资源收集、保存和良种选育提供依据。
1 材料与方法
1.1 材料采集
依据巨柏地理分布并通过实地调查,确定了此次种源的主要采集区域。供试巨柏种子于2020年9月分别采集于林芝种群、米林种群和朗县种群[14]。林芝种群巨柏集中分布于尼洋河流域巴结乡附近的半阳坡坡地上,海拔3 040~3 250 m,年均降水量688 mm,年均温8.7 ℃,土壤沙质;米林种群巨柏集中分布于雅鲁藏布江北岸朗县下游170 km处的石滩地及阳坡中下部高山松林与灌丛之间,海拔2 900~3 100 m,年均降水量700 mm,年均温8.2 ℃,河谷地带风沙地貌明显;朗县种群巨柏主要带状分布在砾石河滩地及河谷两侧的阳坡和半阳坡沙地上,海拔2 900~3 550 m,年均降水量420 mm,年均温8.9 ℃,土壤沙质化。3个地理分布区域气候、土壤、地形等生态因子差异持续影响着巨柏种群及群落特征变化[15-16]。
1.2 种子形态特征测定
在每个巨柏种群内分别选择了100粒种子用作性状测定。用电子游标卡尺测定其长度(纵轴为长,mm)、宽度(腹面横向最宽处为其宽,mm)和高度(种子平放时上下腹面间距离为高,mm),精确到0.01 mm,并计算标准差(SD)、变异系数(CV)和种形指数(EI)[17];随机选取不同种群巨柏种子各1 000粒,用电子天平称量千粒重,重复3次。
1.3 种子萌发试验
巨柏种子发芽试验于翌年4月进行,随机选取3个种群的巨柏种子,利用恒温培养箱进行培养。种子置于培养皿内,润湿的细沙为基质,设置25~30 ℃变温,模拟自然光照,胚根长至0.5 cm左右为发芽标准,每个处理100粒种子,3个重复。随时观察发芽情况,分别对3个种群种子的初始发芽日期、发芽粒数和发芽天数进行统计,后期连续5 d无新种子发芽视为萌发结束。根据发芽高峰日发芽的种子数与供试种子总数的比来计算发芽势(%)。发芽种子总数与供试种子总数的比来计算发芽率(%)[18]。
1.4 数据分析
使用Excel 2019软件进行数据处理和图表制作,用SPSS 25.0软件进行单因素方差分析和显著性检验。
2 结果与分析
2.1 种子表型变异特征
由表1可知,3个种群内巨柏种子长度平均为4.09 mm,变化范围为3.68~3.82 mm;林芝和朗县种群的种子平均长度一样,米林种群种子平均长度最小;种子宽度的平均值为3.14 mm,变化范围为2.71~3.45 mm,其中来自米林种群的平均种宽最大,来自林芝种群的平均种宽最小。种形指数平均值为1.30,变化范围为1.28~1.35,其中朗县种群的平均种形指数最大,林芝种群的平均种形指数最小。米林种群的种子高度最大,林芝种群次之,朗县种群最小;3个种群巨柏种子大小差异不大,千粒重变化范围为3.12~4.18 g,平均值为3.65 g,其中米林种群的平均千粒重最大,朗县种群的平均千粒重最小。质量指标的变化幅度不大。从种子性状指标间的变异系数来看,种长、种宽、种高、种形指数、千粒重分别为9.03%、5.43%、9.80%、25.00%和 8.80%,种子性状的变异系数越大,离散程度越大,性状指标的稳定性就越差。3个种群内种形指数变化幅度最大,种长次之,种宽差异最小。3个巨柏种群内5项表型指标变异程度存在显著性差异,种形指数的变异系数显著高于种长、种宽、种高和千粒重,这表明巨柏种子形状指标受地理分布区域环境影响较大;种长指标变异系数次之,亦受地理环境影响表现出不稳定性。
进行变化方差分析且3种群间两两比较结果显示(如图1),3个种群巨柏种子的种长、种宽、种高、种形指数和千粒重都表现出不同的差异性。朗县种群的种子最长,高于林芝和米林种群的种子,但差异不显著;米林种群的种子宽最大,朗县种群次之,林芝种群最小,米林和林芝种群间差异显著;林芝种群的种子高度最大,朗县种群的最小,林芝和朗县种群之间差异显著;就种形指数而言,朗县和林芝种群的差异不显著,与米林种群的差异显著;米林种群的千粒重最大,朗县种群的最小,两者间差异显著。种子形状与质量指标受限于地理分布范围内资源或气候梯度变化,也正是对区域生境差异的反映。

表2 3个种群巨柏种子萌发特征Table 2 Seed germination characteristics of three populations of Cupressus gigantea
2.2 种子萌发特征
对林芝、米林、朗县3个种群的巨柏种子萌发特征指标进行统计分析(见表2),林芝和朗县种群的巨柏种子均于第9天开始发芽,米林种群的种子于第10天开始发芽。发芽后期连续5 d无新种子发芽时统计总发芽天数,米林种群的发芽最晚且发芽时间也最短(14 d),林芝种群的发芽总天数15 d,朗县种群持续发芽时间最长(18 d)。随着发芽时间增长,米林和朗县种群巨柏种子的发芽率均呈增加趋势,米林种群种子于第18天达到发芽高峰期,发芽势为4%;朗县种群发芽率增加相对较快,于第21天达到发芽高峰期,发芽势为21%;而林芝种群种子于第9天发芽1粒后,再无发芽迹象。
3个种群巨柏种子的发芽率均很低,但差异显著(如图2)。初始发芽时,朗县种群的种子发芽率就显著高于其他2个种群种子的发芽率;统计至发芽21 d时,3种群种子发芽势、发芽率均达到最大差异。朗县种群种子萌动较快,出芽整齐,发芽率最高(42%),且芽苗粗壮呈红白色,处于健康状态,有利于后期幼苗的成活;出苗状态米林种群次之,林芝种群最差。
3 讨 论
种子是苗木繁育的关键,种子表型特征研究又是优良种源筛选的重要基础性研究。不同种群巨柏种子形态指标存在差异,更明显表现在种形指数和千粒重方面[19]。种形指数最不稳定的是米林种群,其次林芝种群,最小的是朗县种群,由此可见朗县种群种子质量最稳定;虽然朗县种群千粒重最小,但其变异系数也最小,也表明了朗县种群种子性状最稳定性,这可能是朗县种子长、宽、高协同进化的结果。种子表型特征受遗传和环境因素共同调控,巨柏种子质量与结种部位、球果大小等相关[13],但种子表型特征结合地理区域环境影响的研究有待深入。
种子萌发时将储藏的营养物质全部激活提供给幼苗[20],种子质量是基础,种子萌发特征亦是种子质量的评价指标。3个种群巨柏种子发芽率差异显著,林芝巨柏古树群种子发芽率仅为1%,朗县种群发芽率为其42倍;朗县种子大小及千粒重均最小,但种子萌发率却最高,该结果也与3个种群幼苗调查现状相符合。本研究在同样发芽条件下进行的3种群巨柏种子萌发特征探索,不同地理区域气候变化和模拟原生境条件是否会提高其发芽能力有待进一步研究。
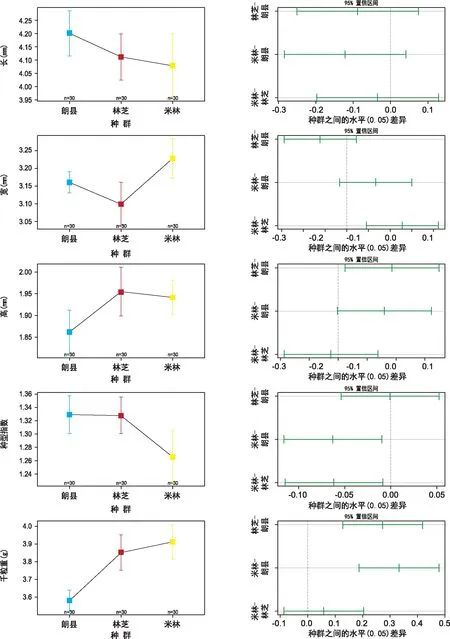
图1 3个种群巨柏种子表型特征方差分析 Fig.1 Variance analysis for seed phenotypic characteristics of Cupressus gigantea from three populations
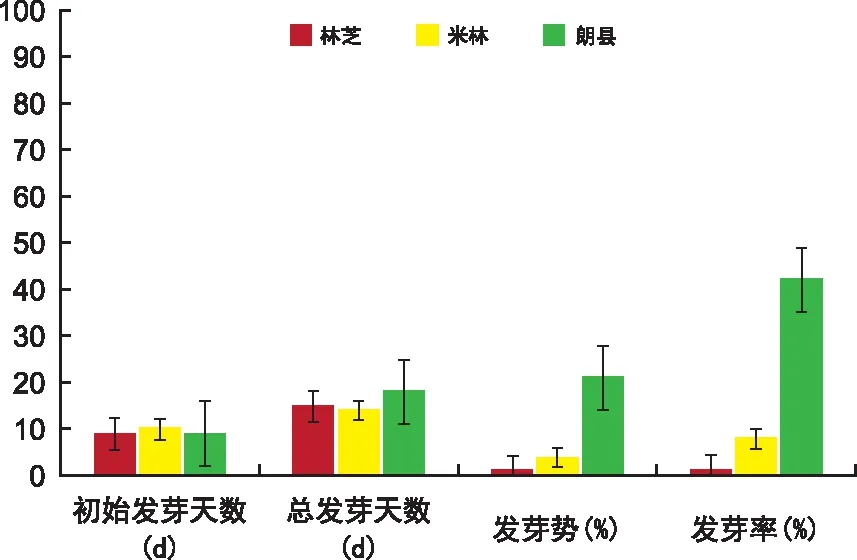
图2 3个种群巨柏种子发芽特征Fig.2 Seed germination characteristics of Cupressus gigantea
本研究分析了3种群巨柏种子表型特征和萌发特性,建议进行巨柏种苗繁育时,选择朗县种群的种源最佳;该研究也仅限于巨柏野生地理区域小尺度范围内,为了选出更稳定的优良种源,今后应该综合地理分布范围内资源或气候变化梯度、种群对地理变异格局的响应策略等因素,就表型和遗传等方面进行多尺度、多层面研究。
