毛棉杜鹃根际与非根际土壤微生物群落多样性*
2022-05-11彭金根龚金玉范玉海张银凤白宇清王艳梅谢利娟
彭金根 龚金玉, 范玉海 张 华 张银凤, 白宇清 王艳梅 谢利娟
(1.深圳职业技术学院建筑工程学院 深圳 518055; 2.河南农业大学林学院 郑州 450002; 3.日照市自然资源和规划局 日照 276827; 4.深圳市梧桐山风景区管理处 深圳 518004)
根际是指受植物根系影响的微型区域,是植物根系与土壤相互作用的主要场所(Shietal., 2015)。在这个微型区域内植物根系、土壤微生物群落以及土壤环境三者相互影响形成有利于植物生长的稳定微环境(黄娜等, 2019; Suretteetal., 2003)。根际微生物是指受植物根系影响最大的微生物群体,其对土壤养分的转化(石柯等, 2019)、植物的生长发育(陆雅海等, 2006)、抗逆性(孙英杰等, 2018)等都具有重要的作用。有研究表明植物根际存在某些有益微生物,它们通过抑制植物病原菌或者分泌有益激素或有机物质来促进植物的生长(王光华等, 2004),而杜鹃花科植物根系能与某些土壤真菌共生形成典型根系形态的共生体,且能增强宿主植物的抗逆性以及养分吸收(李亚东等, 1996; 欧静等, 2012); 同时植物根际也存在一些使植物感病的病原菌(阮柳等, 2016)。侯慧等(2016)对经济作物的研究发现,植物的连作障碍是由于植物在长期的生长过程中,根际微生物群落结构失衡,因此对于植物根系微生物功能及群落结构研究对植物的生长发育很有必要。
毛棉杜鹃(Rhododendronmoulmainense)是杜鹃花科(Ericaceae)杜鹃花属的常绿植物,花期是每年3—4月,花序顶生,花朵簇生,花大色艳,具有极高的景观价值及生态价值(黄滔等, 2010)。梧桐山毛棉杜鹃“十里杜鹃”景观现已形成深圳市特色景观,每年花期吸引成千上万的市民上山赏花。目前毛棉杜鹃的研究已有较多的报道,主要包括组织培养(赵富群等, 2017)、花芽分化(孙敏, 2009)、光适应(白宇清, 2016)以及繁殖生物学(白宇清等, 2017)等。毛棉杜鹃属于浅根系植物,根系的发育对毛棉杜鹃的生长至关重要。廖映辉等(2016)对海南霸王岭毛棉杜鹃根部真菌的研究表明,根部存在丰富的真菌资源,如子囊菌门(Ascomycota)、担子菌门(Basidimycota)等。周玉洁等(2017)研究表明接种福廷瓶头霉菌(Phialocephalafortinii)和聚多曲霉菌(Aspergillussydoeii)可以提高其幼苗的光合能力,促进其植株及根系的生长。本文利用高通量测序技术研究梧桐山毛棉杜鹃根际与非根际土壤微生物群落特征为,探究毛棉杜鹃根际、生态提供依据。
1 材料与方法
1.1 试验材料
2021年7月上旬在深圳梧桐山选取树龄、营养管理水平及长势一致的毛棉杜鹃3块样地,每块样地内保证毛棉杜鹃样树大于5株,在每块样地内利用五点取样法,去除表层的枯枝落叶,然后挖取0~20 cm的毛棉杜鹃细根,抖落附着在根上的大块土壤作为非根际土壤;将附着在根系2 mm范围内的土壤利用无菌刷收集作为根际土;将每块样地内5个点的根际土壤混合均匀作为本样地的根际土壤;本样地内5个点的非根际土壤混合均匀作为该样地的非根际土壤样本。利用无菌袋及车载冰箱将土壤带回实验室进行后续处理,将每份土壤样本分成2份,其中一份过2 mm筛自然阴干后用于测定土壤理化性质; 另一份用干冰冷冻寄送北京百迈客生物科技有限公司以提取土壤微生物。
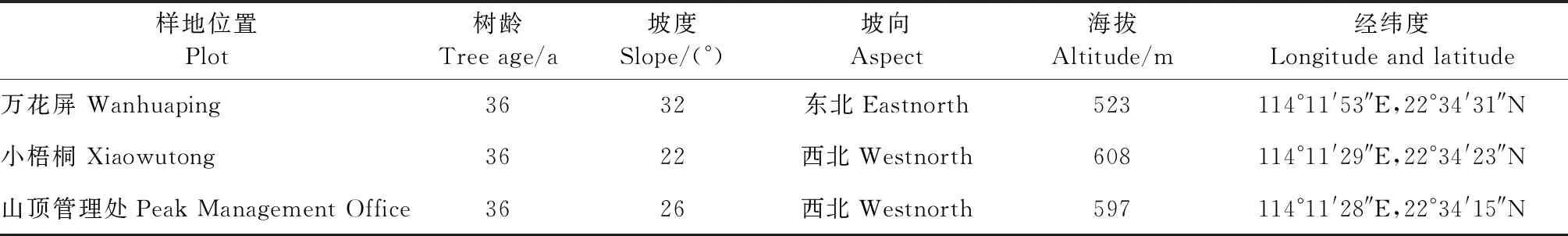
表1 梧桐山样地基本情况
1.2 试验方法
毛棉杜鹃根系土壤理化性质的测定主要参考刘光崧(1996)。土壤pH值的测定利用酸度计法(1∶2.5=土∶ 水); 土壤有机质(SOC)采用重铬酸钾氧化-外加热法; 全氮(TN)的测定利用凯氏定氮法; 全磷(TP)的测定利用其氧化钠熔融-钼锑抗比色法; 全钾(TK)的测定利用其氧化钠熔融-火焰光度计法; 碱解氮(AN)的测定利用碱解扩散法; 速效钾(AK)的测定采用乙酸铵浸提-火焰光度法; 土壤有效磷(AP)的测定采用盐酸-氯化铵浸提法。
1.3 土壤DNA的提取
利用TGuide S96磁珠法土壤基因组试剂盒提取土壤DNA,使用synergy HTX酶标仪对已提取的核酸进行浓度检测,并通过保守区设计引物,再在引物尾端加上测序接头,进行PCR扩增,扩增时用引物ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和 ITS2(5′-GCTGCGTTCTTCATCGATGC-3′)扩增真菌ITS1区; 用338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)扩增细菌v3+v4区,然后对其产物进行纯化、定量和均一化以构建文库,已建成的文库首先采用Qsep-400方法进行文库质量检验,质检合格的文库再用Illumina Novaseq 6000进行检测。高通量测序所得到的原始图像数据文件,经过碱基识别分析后转换为原始序列(sequenced reads),然后对原始序列进行过滤、拼接、去除嵌合体等得到有效序列(effective reads)。
1.4 数据分析
土壤理化性质数据分析采用IBM SPSS Statistics 25软件,根际与非根际间比较采用T-test法; 高通量测序数据下机后得到原始数据,用FLASHv1.2.11、Trimmomatic、UCHIME等工具对原始数据进行拼接、过滤、去除嵌合体,再基于97%的相似性对数据进行聚类,以测序得到所有序列数的0.005%作为阈值过滤后划分OTU,再参考Silva(细菌16S)、Unite(真菌ITS)分类学数据库对OTU进行分类学统计。使用QIIME2软件,对样品的Alpha多样性指数进行评估,采用对样本序列进行随机取样的方法,构建稀释性曲线并统计多样性指数等,比较不同土壤样本的真菌与细菌丰度及多样性; 利用QIIME软件进行 Beta多样性分析; 真菌的功能预测分析利用FUNGuild数据库,置信水平选用可能(probable)和很可能(highly probable)(Nguyenetal., 2016)。通过采用R软件中的Vegan软件包,对土壤理化性质与优势微生物进行RDA分析。
2 结果与分析
2.1 根际与非根际土壤理化性质分析
毛棉杜鹃根际土壤理化性质见表2。根际土壤pH值及速效钾含量显著低于非根际土壤(P<0.05),根际土壤速效磷显著高于非根际土壤(P<0.05)。根际土壤有机质、全氮、碱解氮均高于非根际土壤,但差异不显著。
2.2 微生物多样性及群落结构
2.2.1 细菌与真菌稀释性曲线 以测序深度与物种数目来构建稀释性曲线从已测得的序列中随机抽取OTU序列并统计物种数目。并由图1可知,当细菌OTU数目达到1 200左右,真菌OTU 数目达到1 000左右,稀释性曲线趋于平稳,说明此时测序量已足够覆盖土壤中的微生物类群。
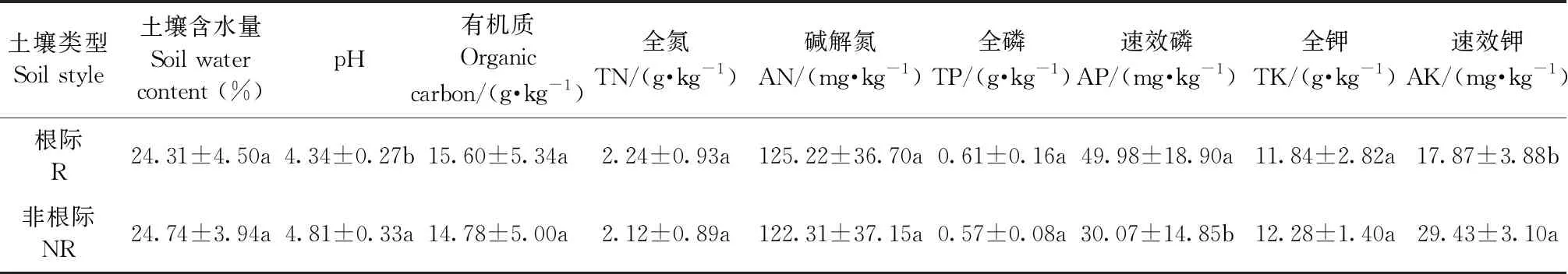
表2 毛棉杜鹃根际与非根际土壤理化性质①
2.2.2 根际与非根际微生物群落丰富度和多样性 基于OTUs分类结果绘制Venn图,由图2A可知,根际与非根际土壤细菌共有OTU为1 382个,其中非根际特有OTUs为144个,占总OTU数目的9%,根际特有OTU为51,占总OTU数目的3%。图2B可知,根际与非根际土壤真菌共有OTU为1 231个,其中非根际特有OTU为10个,根际特有OTU为21个,根际特有OTU数目高于非根际土壤。
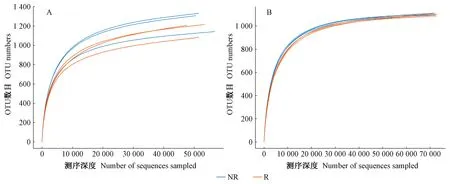
图1 细菌(A)、真菌(B)稀释性曲线

图2 毛棉杜鹃根际和非根际土壤细菌(A)、真菌(B)Venn图
Alpha多样性(Alpha diversity)可反映单个样品的物种丰度(richness)及多样性(diversity),主要包括Chao1指数、ACE指数等。毛棉杜鹃细菌与真菌根际与非根际Alpha多样性指数均无显著性差异(P>0.05),但非根际细菌土壤样本具有更高的微生物多样性和物种丰度,而真菌根际与非根际相差不大(图3A、3B)。毛棉杜鹃非根际土壤细菌与真菌Shannon指数与Chao1指数均高于根际土壤(P>0.05),表明了非根际土壤细菌较真菌具有更高的多样性和丰富度(图3C、3D)。而根际与非根际土壤环境中,细菌相较于真菌具有更高的多样性和丰富度(P<0.05),说明毛棉杜鹃根系环境仍属于细菌主导型,细菌相较于真菌具有更高的物种丰度及多样性。
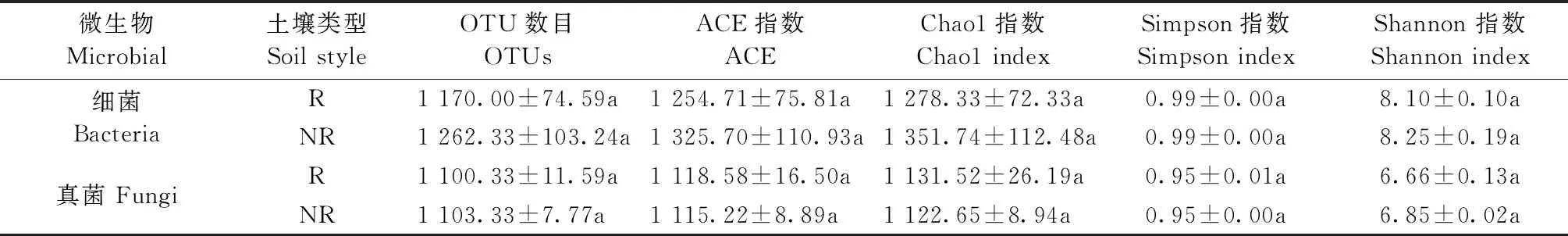
表3 毛棉杜鹃根际与非根际土壤细菌和真菌群落的多样性指数
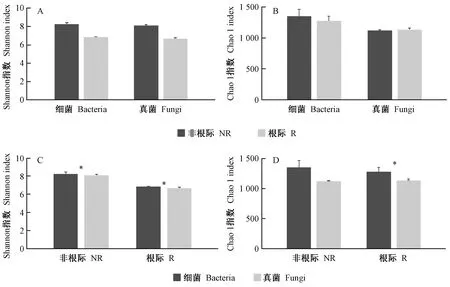
图3 根际与非根际土壤细菌和真菌群落的多样性指数
2.2.3 根际与非根际土壤微生物组成 将原始序列过滤后统计出各样品中各等级的物种数。以Silva、UNITE数据库为参考,可获得真菌、细菌物种统计表。由表3可知,毛棉杜鹃根际与非根际土壤微生物多样性丰富,其中根际细菌分别注释到23门、64纲、125个目、188科、277个属以及291种,非根际注释到25门、69纲、134目、201科、291属以及305种,非根际在细菌水平上多样性是稍高于根际土壤; 而根际真菌注释到8门、29纲、77目、180科、352 属、520种,非根际真菌注释到8门、29纲、77目、180科、353属、518种,根际与非根际真菌多样性相差不大。

表3 根际、非根际土壤细菌、真菌物种统计
由门水平物种丰度图4A可知,毛棉杜鹃根际土壤优势细菌门前10名分别为酸杆菌门(Acidobacteria)42.47%、变形菌门(Proteobacteria)30.72%、放线菌门(Actinobacteria)6.80%、绿弯菌门(Chloroflexi)4.93%、浮霉菌门(Planctomycetes)4.42%、疣微杆菌门(Verrucomicrobia)2.84%、WPS-2 4.58%、芽单胞菌门(Gemmatimonadetes)0.87%、拟杆菌门(Bacteroidetes)0.93%以及厚壁菌门(Firmicutes)0.25%。非根际土壤优势细菌门主要是酸杆菌门46.33%、变形菌门26.26%、放线菌门6.38%,其中非根际土壤样本中酸杆菌门占比比根际土壤样本高9.1%,而根际土壤样本中变形菌门占比比非根际样本高17%。
毛棉杜鹃根际土壤真菌优势物种菌门共注释到8个门如图4B,其中优势真菌门为子囊菌门和担子菌门,两者占比高达90%以上。而根际与非根际土壤优势真菌门分别是子囊菌门64.95%、担子菌门29.51%、被孢菌门(Mortierellomycota)1%、罗兹菌门(Rozellomycota)0.81%、壶菌门(Chytridiomycota)0.34%、油壶菌门(Olpidiomycota)0.1%、球囊菌门(Glomeromycota)0.05%。非根际土壤优势菌门为子囊菌门72.21%、担子菌门22.05%、被孢菌门1.26%。由图4B可知,非根际土壤子囊菌门丰度比根际土壤高11.2%,而根际土壤担子菌门丰度比非根际高33.8%。
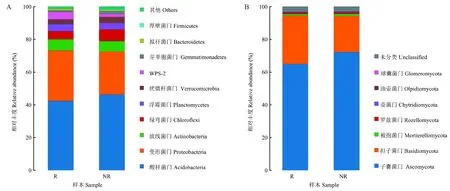
图4 根际与非根际土壤细菌、真菌门水平物种分布
由目水平物种丰度图可知,毛棉杜鹃根际与非根际土壤优势细菌目(图5A),主要是为 Subgroup_2、酸杆菌目(Acidobacteriales)、Solibacterales、根瘤菌目(Rhizobiales)等。根际与非根际土壤优势真菌目(图5B),主要是散囊菌目(Eurotiales)、肉座菌目(Hypocreales)、蜡壳耳目(Sebacinales)等。
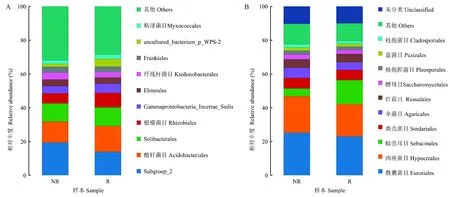
图5 根际与非根际土壤细菌、真菌目水平物种分布
2.2.4 根际与非根际土壤β多样性分析 主成分分析(PCA)可以将根际与非根际土壤样品差异反映在二维坐标轴上,样本在坐标轴上的距离越小,表示2个样本的组成越相似。主坐标轴显示细菌在根际土壤差异较小,在非根际土壤差异较大(图6A); 真菌β多样性在非根际土壤中分布较为集中,差异较小,在根际土壤差异要高于非根际土壤(图6B)。
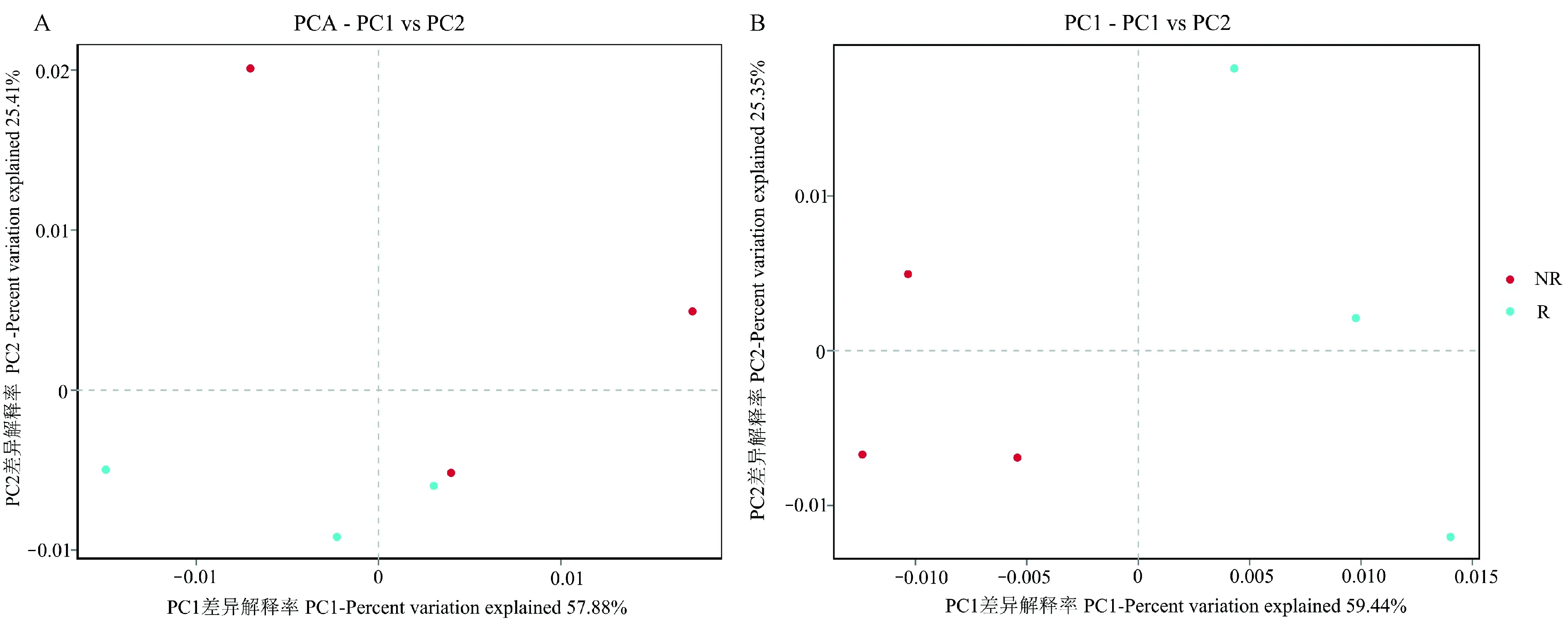
图6 毛棉杜鹃根际与非根际土壤细菌(A)、真菌(B)主坐标分析
2.2.5 根际与非根际土壤真菌功能基因预测 通过FUNGuild对毛棉杜鹃根际与非根际土壤真菌进行功能预测分析,将置信水平为Highly Probable和Probable的物种筛选出来并进行可视化,从而得到真菌生态营养类型及各营养类型在不同样本中的占比情况。Trophic Mode结果显示,毛棉杜鹃根际与非根际土壤真菌类群主要预测得到7种营养类型(图7A),分别是共生营养型(symbiotroph)、腐生-共生营养型(saprotroph-symbiotroph)、腐生营养型(saprotroph)、病理-共生营养型(pathotroph-symbiotroph)、病理-腐生-共生营养型(pathotroph-saprotroph-symbiotroph)、病理-腐生营养型(pathotroph-saprotroph)和病理营养型(pathotroph)。根际与非根际占比最高的营养型均为病理-腐生营养型、共生营养型和腐生营养型。其中,根际与非根际主要营养型为共生营养型、腐生营养型、病理-腐生营养型,三者丰度之和高达80%以上。根际共生营养型真菌丰度比非根际高42%,说明共生营养型真菌在根际富集,主要序列为OTU7、OTU10、OTU22等,其注释为担子菌门的红菇属Russula等。
各营养型丰度占比情况见表4。根际共生营养型真菌丰度比非根际上升42%,其余营养型真菌均为非根际高于根际。基于Guild结果根际与非根际预测共得到24个生态共位群,其相对丰度如图7B所示,其中非根际(NR)优势共位群分别是木腐生菌(wood saprotroph,22%),植物病原菌(plant pathogen,21%)和动物病原菌(animal pathogen, 20%)等共位群。根际优势生态位则为木腐生菌(wood saprotroph,21%),主要分类单元注释为小柱孢属(Scytalidium)。植物病原菌(plant pathogen, 20%),主要分类单元注释为粉红螺旋聚孢霉属(Clonostachys)以及动物病原菌(animal pathogen, 19%),主要分类单元注释为蜡蚧轮枝菌属(Lecanicillium)等。

表4 根际与非根际土壤真菌营养类型及丰度
2.2.6 毛棉杜鹃土壤优势微生物的影响因子 对毛棉杜鹃根际与非根际土壤微生物及其土壤理化性质进行冗余分析,结果见图8。对于土壤细菌群落RDA两轴的贡献率分别为36.56%和29.01%,所选5个土壤基本性质共解释65%的总特征值,其中有机质,碱解氮与浮霉菌门、疣微杆菌门等呈正相关。pH值、速效钾与酸杆菌门、绿弯菌门以及厚壁菌门呈正相关。对于土壤真菌群落RDA分析,两轴的贡献率分别为45.62%、28.74%,总贡献率达74.3%,其中有机质及碱解氮含量与罗兹菌门、壶菌门之间呈正相关。pH值以及速效钾与子囊菌门、油壶菌门以及被孢菌门之间呈正相关。
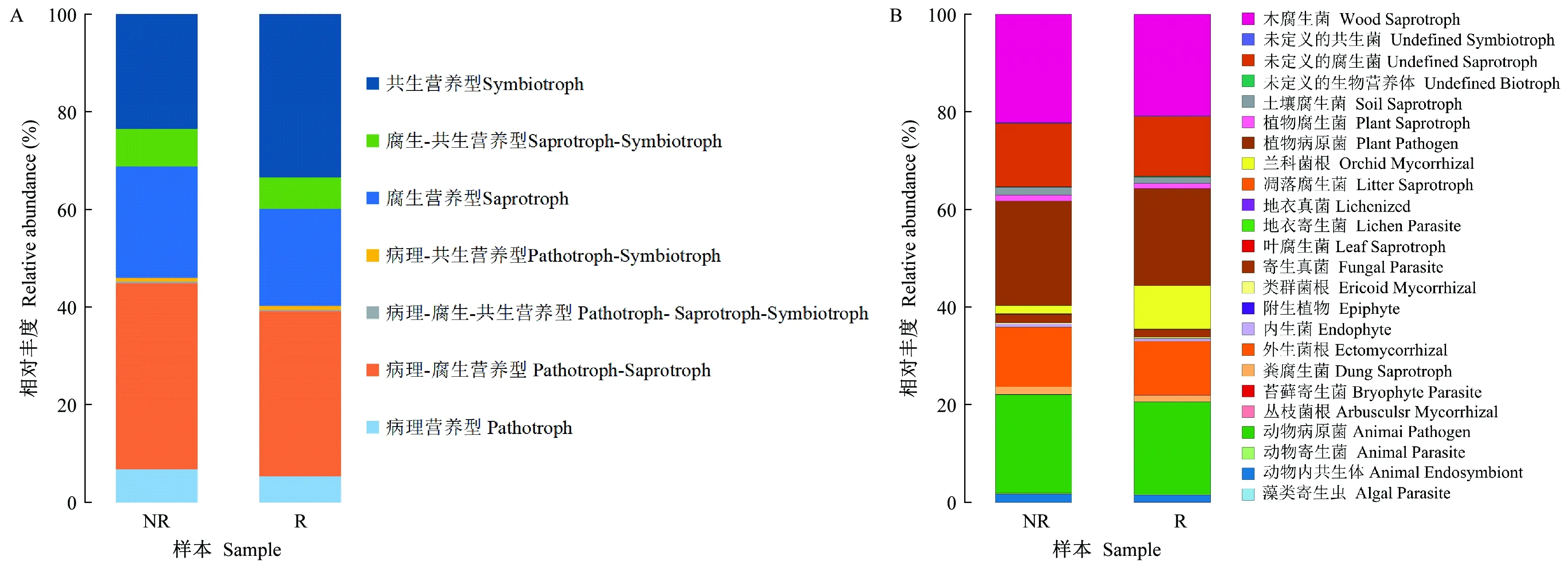
图7 根际与非根际土壤真菌功能基因预测
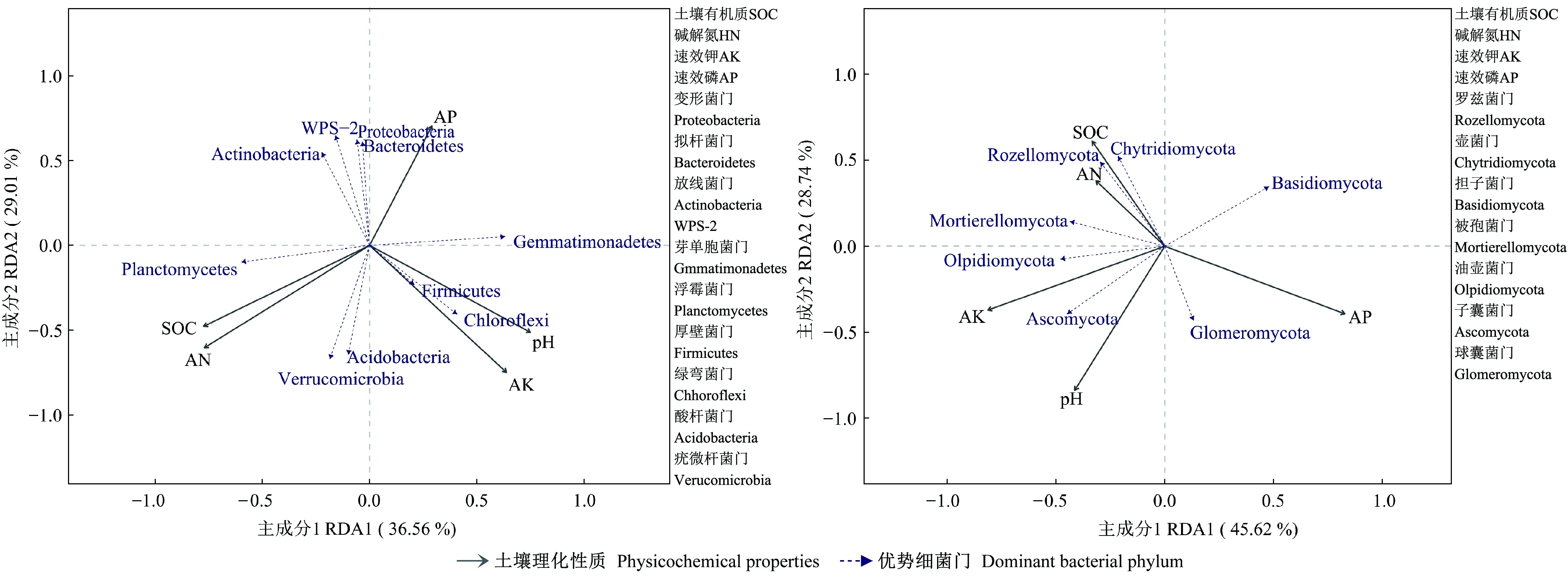
图8 优势细菌(A)、真菌(B)与土壤理化性质冗余分析
3 讨论
土壤理化性质对植物的生长、发育至关重要,毛棉杜鹃根际土壤pH值明显低于非根际土壤,出现这种现象的原因可能是由于毛棉杜鹃根系产生的分泌物进入根际土壤,从而引起根际土壤pH值降低。根际有机质、全氮、碱解氮等营养元素含量均高于非根际,其原因可能是根系分泌的有机物等进入根际土壤,从而使根际与非根际土壤在理化性质等方面产生差别。而土壤中营养物质在根际区域发生富集现象在七叶一枝花(Parispolyphylla)(郑梅霞等, 2020)、南酸枣(Choerospondiasaxillaris)(童琪等, 2019)、华北落叶松(Larixprincipis-rupprechtii)(郭辉等, 2020)等植物中均已被证实。
土壤微生物是土壤中重要的组成部分,其多样性及丰富度可以间接的反映土壤质量及土壤肥力(Wangetal., 2019)。已有研究表明,由于植物的根系分泌以及淋溶等活动,导致植物根际与非根际在土壤理化性质以及微生物群落结构等方面存在差异(Mousavietal., 2018)。Yang等(2010)研究表明,土壤微生物主要以细菌为主,本研究结果中同样也是细菌种类高于真菌,但真菌种类也相对较高。李岩等(2018)对不同生境黑果枸杞(Lyciumruthenicum)根际与非根际土壤微生物的研究表明,根际真菌微生物群落结构多样性低于非根际,与本文研究结果相同,但在本研究中非根际细菌高于根际,出现这种现象的原因很有可能是深圳夏季多雨,而高温高湿的土壤环境有利于微生物的增殖,在特定的气候条件下根系分泌物的选择功能,也会使根际微生物多样性低于非根际,即形成根际负效应; 通过FUNGuild分析表明,土壤真菌类群病理-腐生营养型和病理营养型等功能类群。因此,根际的负效应有可能是毛棉杜鹃根际土壤真菌多为致病菌类型,导致植物根系分泌抑制物质对病原菌产生抵抗,进而导致根际微生物多样性低于非根际的现象。马志良等(2019)研究发现,增温对高寒灌丛土壤根际与非根际微生物的影响不同,且在不同季节也会产生不同的根际效应。蒋治岩等(2021)在研究表明,植物的根际效应与根际形成的菌根类型以及植物种类均有关系。
毛棉杜鹃属典型的杜鹃花科植物,土壤中某些真菌可以侵染其植物根系并与之形成杜鹃花类菌根(ericoid mycorrhizas,ERM)(张艳华等, 2021),植物菌根的形成在促进宿主营养物质吸收、增强抗逆性等方面具有重要作用。郑钰等(2010)在银叶杜鹃(Rhododendronargyrophyllum)和繁花杜鹃(R.floribundum)根部真菌研究中发现,杜鹃属植物具有丰富的真菌资源,蜡壳耳目和柔膜菌目(Helotiales)是形成其菌根的主要真菌。唐燕等(2019)对云南轿子山腋花杜鹃(R.racemosum)菌根的研究发现,瑞士无梗囊霉(Acaulosporarehmii)、布氏球囊霉(Glomusbrohultii)等与腋花杜鹃根系形成丛枝菌根(arbuscular mycorrhizas,AM)。廖映辉等(2016)对毛棉杜鹃根部真菌的研究发现,其具有丰富的根部真菌资源,主要是子囊菌门、担子菌门等,与本文的研究结果一致。真菌群落结构与细菌相比更加复杂,有可能是为了适应恶劣的生存环境,从而进化形成具有多种营养方式,这是真菌群落为了适应恶劣环境而形成的较为先进的生存结构(熊丹等, 2020)。土壤中病理真菌的大量繁殖会导致植物患病,严重的会导致植物体的死亡(姜子德等, 2000),任伟恒等(2020)对马缨杜鹃(R.delavayi)的研究发现,感染根腐病的杜鹃根际真菌明显增多。
土壤理化性质与优势微生物的冗余分析结果表明,土壤pH值、有机质、碱解氮、有效磷、有效钾含量均对土壤优势微生物有不同影响,有机质,碱解氮与浮霉菌门、疣微杆菌门等呈正相关,pH值、速效钾与酸杆菌门、绿弯菌门以及厚壁菌门呈正相关。字洪标等(2017)对青海省不同林分微生物群落结构研究发现,土壤pH值是影响微生物群落的主要环境因子,有机质及碱解氮含量与罗兹菌门、壶菌门之间呈正相关。土壤pH值以及速效钾与子囊菌门、油壶菌门、被孢菌门之间呈正相关。
4 结论
梧桐山毛棉杜鹃根际与非根际土壤理化性质除土壤pH值、速效钾和全钾含量外,均为根际土壤高于非根际,土壤pH值、速效钾含量显著性低于非根际。根际与非根际优势细菌类群相同,均为酸杆菌门、变形菌门、放线菌门等。根际与非根际优势真菌为子囊菌门、担子菌门、被孢菌门等。毛棉杜鹃根际与非根际优势细菌目,主要是Subgroup_2、酸杆菌目、Solibacterales、根瘤菌目等。根际与非根际优势真菌目主要是散囊菌目、肉座菌目、蜡壳耳目等。毛棉杜鹃根系真菌主要是病理-腐生营养型、共生营养型等,且根际与非根际在共生营养型真菌丰度具有较大差异,毛棉杜鹃根际含有丰富的共生真菌资源,且根际与非根际的真菌功能结构有差异。冗余分析结果表明,土壤有机质,碱解氮与浮霉菌门、疣微杆菌门等细菌门呈正相关; 土壤pH值、速效钾与酸杆菌门、绿弯菌门以及厚壁菌门呈正相关。有机质及碱解氮含量与罗兹菌门、壶菌门等真菌门呈正相关; 土壤pH值以及速效钾与子囊菌门、油壶菌门以及被孢菌门等真菌门呈正相关。
