长期不同养分缺乏对冬油菜土壤微生物群落组成及多样性的影响
2022-06-14甘国渝陈佛文邹家龙高旭园李继福
甘国渝,陈佛文,邹家龙,何 洁,陈 伟,高旭园,李继福*
(1.湿地生态与农业利用教育部工程研究中心/长江大学农学院,湖北 荆州 434025;2.荆州市荆州区农业技术推广中心,湖北 荆州 434025)
江汉平原是湖北省冬油菜的主产区,常年种植规模约67万hm2,占全省播种面积的60%以上[1]。冬油菜作为南方越冬作物,基本不与主粮争地,且有养地功能,这是因为油菜秸秆和落叶还田可补充土壤有机质、提高土壤肥力和改善土壤生物学特性,如微生物组成及其多样性[2]。长期冬油菜种植对土壤微生物群落产生选择和富集的作用,而微生物又是土壤的重要组成部分,在土壤有机碳氮矿化和矿质元素地球化学循环中起着重要作用,是衡量土壤质量的重要生物指标[3]。施肥作为油菜栽培过程中的重要环节,对产量形成和养分供给产生显著影响,而施肥或者肥料投入不足会导致根际微生物的优势种群和相对含量发生变化,从而影响有机物矿化进程和养分供给水平[4]。因此,开展长期养分缺乏胁迫下土壤微生物组成和多样性研究有助于从微生物层面解析冬油菜根际微生态环境变化对土壤生物肥力改变的反馈机制。土壤微生物作为土壤养分的补充储备库,可为冬油菜生长提供必需的营养物质。冬油菜根际微生物紧密依附在根际土壤颗粒中,与根系存在适应协同进化的关系。根际微生物能够释放营养物质到土壤介质中,促进作物成长[5],而植物通过根系分泌物可为微生物繁殖提供碳源。有研究发现,化肥平衡施用、有机无机肥配施均显著提高土壤中细菌数量、多样性和丰富度,有机无机肥配施还改变细菌群落结构,特别是降低放线菌门、增加拟杆菌门的相对丰度,更有利于土壤生态系统的稳定和健康,而肥料投入不足则会导致土壤肥力下降和生态环境失衡[6]。
近20年来,分子生物技术在生命科学领域快速发展,尤其是二代,甚至三代高通量测序的应用,为深入了解根际营养变化对土壤微生物生命发育演化和种群组成的影响提供技术支撑[7-9]。许艳蕊等[10]对玉米根际微生物群的季节性变化研究发现,在营养生长阶段,根际微生物活性和优势物种相对丰度明显高于土壤本体;但在生殖生长阶段,根系活力和可溶性分泌物释放量降低,根际效应也随之消失,导致可培养细菌的生物多样性也明显下降[11-12]。张秫华等[13]利用高通量测序技术分析了川芎根际与非根际土壤真菌群落结构及差异,发现真菌群落均匀度和多样性表现为非根际土>根际土,丰富度表现为根际土>非根际土。另有研究表明,苜蓿根际和非根际土壤细菌、真菌数量均随播量的增加呈先升高后降低的趋势,在播量达到一定值时,根际和非根际土壤微生物数量最大,且根际高于非根际[14]。油菜属于对养分较为敏感的田间作物,尤其对氮肥和磷肥,而油菜根际和非根际微生物对养分胁迫的响应是否存在差异还缺乏足够证据,需要进一步证实。
当前,不同施肥措施对农田细菌和真菌组成及其多样性的研究较为广泛和深入,然而,大量肥料的投入使得土壤积累的养分含量偏高,不仅造成资源的浪费,还影响肥料的吸收利用率[15]。因此,对化肥减施或不施对土壤生物特性的研究还较为薄弱,尤其是在长期单一肥料缺乏下,根际微生物如何适应土壤养分状况还不明确。综合上述问题,本文基于田间肥料定位试验来探明冬油菜根际、非根际细菌和真菌群落组成结构差异及其演化对氮肥、磷肥和钾肥缺乏的响应,为冬油菜根际生态过程中的养分调控、环境污染防治和农业可持续发展提供参考。
1 材料与方法
1.1 试验设计
试验点位于湖北省荆州市荆州区川店镇(30°33′24″ N,112°4′56″ E,海拔52 m),为河流冲积物发育形成的水稻土。2011年水稻季开始进行肥效田间试验,采用一年两熟的水稻-油菜轮作模式。试验前,耕层(0~20 cm)土壤基本理化性质为pH 6.4、全氮0.6 g/kg、碱解氮275.3 mg/kg、有机质28.8 g/kg、有效磷4.44 mg/kg、速效钾96.6 mg/kg、缓效钾528.2 mg/kg。本研究共选取其中4个处理,分别为:(1)施磷钾肥不施氮肥(-N)、(2)施氮钾肥不施磷肥(-P)、(3)施氮磷肥不施钾肥(-K)和(4)施氮磷钾肥(NPK)。各处理重复3次,小区面积20 m2,随机区组排列。各处理除不施用的肥料外,每季肥料用量均按当地推荐, 即:N 180 kg/hm2、P2O590 kg/hm2和K2O 120 kg/hm2,其中油菜季补施硼肥15 kg/hm2。肥料种类为尿素(N 46%)、过磷酸钙(P2O512%)、氯化钾(K2O 60%)和硼砂(B 11%)。具体用量和施用方法为:水稻季氮肥分3次施用,其中基肥占50%、分蘖肥和穗肥各占25%;磷肥和钾肥在水稻移栽前作基肥一次性施用。冬油菜季氮肥分3次施用,其中基肥占60%,越冬肥和蕾薹肥分别占20%;磷肥、钾肥和硼肥在油菜移栽前一次性基施。其他田间管理均按当地最优方式进行。
1.2 样品采样与测定
1.2.1 土壤样品
基础土样于2011年5月冬油菜收获后采集,取0~20 cm耕层土壤,按照四分法取1 kg带回实验室,风干磨细过0.85 mm筛,保存。土壤基本理化性质如pH、全氮含量等指标按照常规方法[16]测定。2020年4月,成熟期在各小区选取长势均匀的油菜4株连根拔起,抖掉附着的松散土,用刀片采集根际土壤,作混合样[17]。同时,用不锈钢土钻采集油菜间隙土壤作非根际土,具体标注见表1。
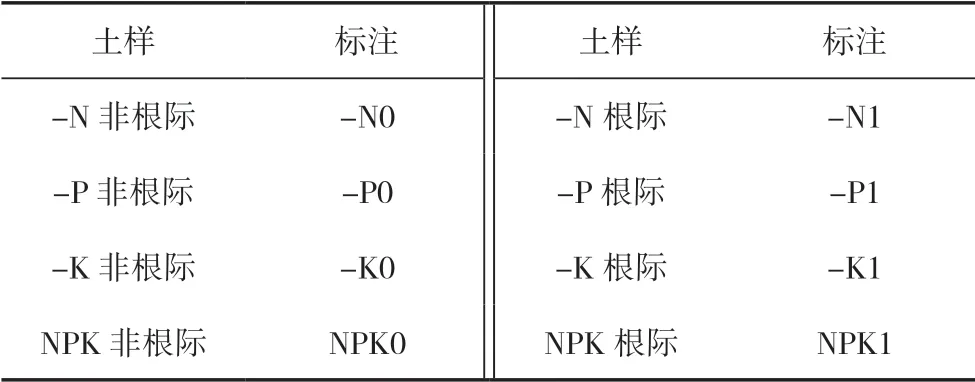
表1 样品名称标注
1.2.2 土壤微生物基因组DNA提取
用强力DNA分离试剂盒(MoBio Laboratories,Carlsbad,CA,USA)提取土壤DNA。对样品进行1.2%琼脂糖凝胶检测DNA的长度和完整性,利用NanoDrop紫外分光光度计(Thermo Scientific,NC2000)检测DNA的浓度和纯度。
1.2.3 DNA扩增及测序
细菌16S rDNA V3~V4区扩增采用前引物序列338F(5’-ACTCCTACGGGAGGCAGCA -3’)和后引物序列806R(5’-GGACTACHVGGGTWTCTAAT-3’)[18],真菌ITS扩增采用前引物序列ITS1F(5’-GGAAGTAAAAGTCGTAACAAGG-3’)和后引物序列ITS2(5’- GCTGCGTTCTTCATCGATGC-3’)[19]。目标条带胶回收纯化后进行文库构建,利用Illumina-MiSeqPE250平台进行高通量测序。
1.3 数据处理
测序原始数据去除引物接头序列和各低质量碱基后进行拼接,舍弃长度短于160 bp的序列,去除非特异性扩增序列及嵌合体后,得到每个样本的有效序列数据。以97%为规定阈值,对16S和18S序列划分可操作分类单元(OTU)。采用QIIME,调用UCLUST序列对比工具按97%的序列相似度进行归并,并选取每个OTU度最高的序列作为该OTU的代表序列。针对细菌16S rRNA和真菌18S rRNA分别采用Greengenes数据库和Silva数据库作为OTU分类地位鉴定的模板序列。
试验数据采用Excel 2016、OriginPro 2019b进行计算和处理,采用LSD法检验0.05水平上的差异显著性。每个处理进行3次生物学重复,处理标注见表1。利用R的vegan工具包以微生物群落结构作为响应变量、土壤养分含量作为解释变量进行冗余分析。
2 结果与分析
2.1 冬油菜根际与非根际土壤养分含量
由表2可知,长期养分缺乏导致处理间pH和养分含量有一定的差异。各处理根际养分含量与非根际相比有明显不同。土壤pH结果显示,与非根际相比,各处理根际土壤pH呈下降的趋势,而有机质、有效磷和速效钾含量有富集的趋势,其中-N1、-P1、-K1和NPK1土样的有机质含量比非根际分别提高6.8%、6.7%、6.6%和6.5%;而缓效钾根际非根际之间差异并不显著,这可能与背景值较高有关。
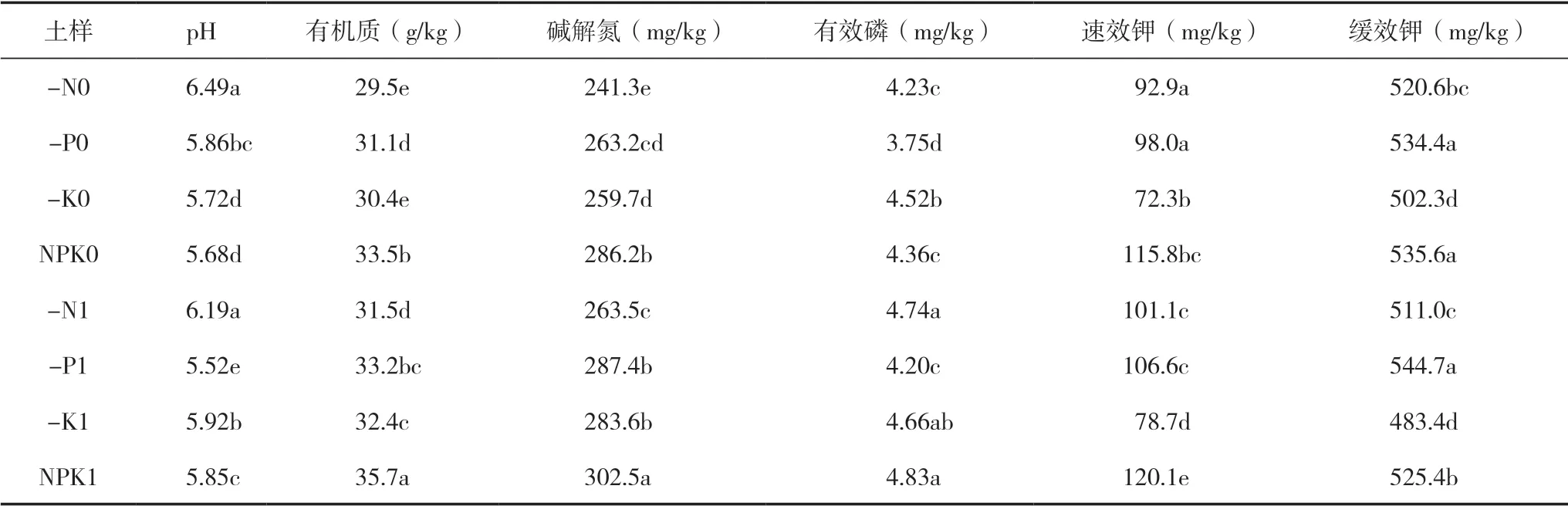
表2 各处理冬油菜根际与非根际土壤养分含量
2.2 冬油菜根际与非根际微生物多样性
配对t检验表明,除-P处理外,各处理油菜根际、非根际的细菌群落多样性(Simpson指数和Shannon指数)无显著性差异(P>0.05),但从趋势上来看,-P和-K处理的非根际土壤细菌群落多样性比根际高,而-N和NPK处理的根际细菌多样性略高于非根际土壤(表3)。与细菌多样性不同,-N、-P处理的油菜根际真菌多样性显著低于非根际(P<0.05),而NPK处理的根际与非根际真菌多样性无明显差异。同时,从Chao1和ACE指数(表3)来看,各处理根际和非根际细菌、真菌的丰富度表现不一。-N处理的根际细菌和真菌丰富度高于非根际,而-P、-K和NPK处理的微生物丰富度则与-N处理相反。总之,长期养分缺乏对冬油菜根际和非根际细菌多样性影响不明显,而对真菌多样性的影响最为显著。

表3 土壤细菌、真菌的丰度和群落结构多样性
2.3 冬油菜根际与非根际微生物群落组成
2.3.1 细菌和真菌门水平分类
由图1A结果可知,细菌门分类水平上,各处理非根际优势物种是绿弯菌门(Chloroflexi)、酸杆菌门(Acidobacteria)、变形菌门(Proteobacteria)和Rokubacteria,其平均相对丰度分别为29.8%、29.0%、15.9%和4.4%;而根际优势物种依次是绿弯菌门、酸杆菌门、变形菌门和Patescibacteria,其平均相对丰度分别为32.0%、22.3%、19.2%和6.4%。具体来看,-P处理根际Patescibacteria的相对丰度显著高于非根际。-N处理下,根际绿弯菌门相对丰度明显高于非根际。NPK处理根际变形菌门相对丰度显著高于非根际,而-K处理的根际与非根际各门细菌相对丰度差异不显著。
图1B结果显示,各处理真菌群落中,子囊菌门(Ascomycota)和Unidentified的相对丰度最高,表明门水平上未被分类的菌群数量仍较多,其中子囊菌门在非根际和根际的平均相对丰度分别为46.4%和57.5%。-N和-K处理下,Rozellomycota在非根际的相对丰度分别可达10.4%和8.0%,在根际的相对丰度分别为10.7%和14.9%。-P和NPK处理下,Rozellomycota门在非根际的平均相对丰度分别为14.3%和17.5%,在根际的平均相对丰度则分别降至3.3%和11.7%。

图1 细菌和真菌门水平上的相对丰度
2.3.2 细菌和真菌属水平分类
细菌属分类水平上(图2A),各处理菌群相对丰度均较低,Uncultured、Unidentified和Other在根际和非根际的相对丰度均较高,表明各处理中不能培养、没有鉴定或者未被分类的菌群较多。优势属种在各处理的表现存在一定的差异,-N、-P和-K处理中,Haliangium在非根际土壤的相对丰度分别为1.4%、1.0%和1.2%,而在根际的相对丰度则分别降至0.5%、0.6%和1.0%。-N和-P处理下,Solibacter在非根际的相对丰度分别为1.2%和1.5%,而在根际土壤的相对丰度分别为1.0%和1.1%。-N和-P处理下,Anaeromyxobacter属在非根际的相对丰度分别为1.0%和0.9%,在根际的相对丰度则分别降至0.3%和0.8%。-K和NPK处理下,Anaeromyxobacter在根际的相对丰度分别为0.9%和2.4%,在非根际的相对丰度则降至0.7%和1.0%。此 外,-N和-P处 理 下,Haliangium、Solibacter和Anaeromyxobacter在非根际的相对丰度均高于根际。

图2 细菌和真菌属水平上的相对丰度
图2B结果显示,除已知物种外,真菌属群落中存在较多的稀有物种(Other),它们相对丰度低,而种类多。-N处理油菜非根际优势类群是Acaulium,相对丰度可达56.2%。支顶孢属(Acremonium)为-P处理油菜非根际优势类群,其相对丰度为17.8%;而Phialocephala和Akanthomvces为-P处理根际优势类群,其相对丰度分别为32.6%和23.2%。油壶菌属(Olpidium)为-K处理根际和NPK处理非根际优势类群,其相对丰度分别为25.3%和21.0%。
2.4 冬油菜根际与非根际微生物群落结构差异
PCoA主成分分析可将样品微生物多样性差异反映在二维坐标图上,主成分轴用来解释差异。图3结果显示,长期不同养分缺乏对油菜根际和非根际微生物差异的影响不一。对细菌而言,第1主成分(PCoA1)和第2主成分(PCoA2)分别解释差异的42.0%和33.1%,合计贡献率占75.1%,表明养分差异使得各处理细菌群落发生较大变异,尤其是-N处理的根际与非根际细菌群落结构变异最大,其他处理根际和非根际细菌群落结构差异也较为明显。对于真菌而言,第1主成分和第2主成分分别解释差异的34.6%和26.3%,合计贡献率占60.9%,低于细菌变异解释率。从图3B可知,-N处理根际与非根际真菌群落分布差距最大,而-P、-K和NPK处理的根际与非根际真菌群落差距较小,受养分缺乏影响不显著。

图3 细菌和真菌PCoA主坐标分析
热图可在属水平上将不同相对丰度的OTU聚类,并根据颜色深浅反映不同处理的菌群物种关系。从图4A可以看出,施肥处理和根际效应显著影响各样本中优势细菌菌群分布和相对丰度。NPK处理根际土壤中,大部分细菌群落的相对丰度较高,而在非根际土中则较低。非根际中的Anaeromyxobacter、Geobacter、Nitrospira、Syntrophorhabdus和Sh765B-TzT-35在-N、-P和NPK处理中的相对丰度较高,而在-K处理中较低。Sh765B-TzT-35和Acidibacter、Gemmatimonas和Gemmatirosa、Gaiella、Ignavibacterium分别为-N、-K、-P、NPK处理的优势属种。根际土壤中Ignavibacterium和Pseudolabrys在-N和-K处理的相对丰度较低,而在-P和NPK处理中则为优势属;Chthonomonas在-P和NPK处理中相对丰度较低,而在-N和-K处理中为优势属。
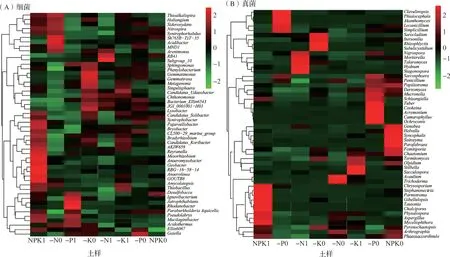
图4 各处理优势细菌和真菌属热图分析
根际真菌Sarocladium、Itersonilia、Sarcos-phaera、Papiliotrema、Mucronella、Saitozyma、Chaetomium和Parmotrema等属以及非根际真菌Olpidium、Physalospora、Myceliophthora和Phaeosaccardinula在4个处理间均具有明显差异(图4B)。此外,-K处理根际中有6种真菌的相对丰度明显高于其他3个处理,包括Derxomyces、Chaetomium、Termitomyces、Olpidium、Stilbella和Sacculospora。-P处理非根际有6种真菌相对丰度明显高于其他3个处理,包括Schizangiella、Tuber、Cookeina、Acremonium、Camarophyllus和Ochroconis。
2.5 冬油菜土壤微生物群落结构与土壤养分冗余分析
冗余分析图可直观地反映土壤养分含量对微生物群落结构的解释程度,影响因素之间的夹角代表各因素之间的相关性大小,其中锐角表示两个因素正相关,直角表示不相关,钝角表示负相关;而射线的长短代表环境因子对研究对象的解释度高低,射线越长,解释度越高,反之越低。由图5可知,土壤速效钾、有机质、碱解氮、有效磷含量及pH是影响土壤细菌(pseudo-F=5.7,P=0.004)和 真 菌(pseudo-F=4.3,P=0.008)群落结构的关键参数,其中pH、速效钾和有效磷含量对细菌群落结构的影响最为显著,而有机质和碱解氮含量对真菌群落结构的影响最为明显。
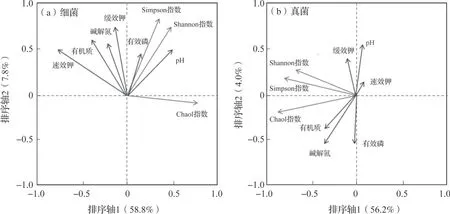
图5 微生物群落结构与土壤养分冗余分析
3 讨论与结论
3.1 长期养分缺乏对冬油菜土壤微生物多样性及养分的影响
Illumina Miseq测序技术具有通量高、灵敏度高、准确性高和运行成本低等特点,近年来已被广泛应用于土壤细菌、古菌和真菌群落组成及其多样性研究。本研究基于Illumina Miseq二代测序技术,分析-N、-P和-K处理对冬油菜根际与非根际土壤细菌和真菌多样性及其群落的组成结构。表2结果显示各处理根际和非根际土壤中细菌的丰富度和均匀度相比真菌较高,这与土壤中微生物以细菌为主有关[20]。-N和NPK处理下,根际细菌多样性比非根际土壤高,而-P和-K处理则相反。4个处理中非根际土壤真菌多样性均比根际土壤高,且长期养分缺乏对细菌和真菌的丰富度影响较为明显。方静等[21]研究表明,合理施用氮肥利于土壤微生物生长和微生物多样性增加,但过量施肥会抑制微生物繁殖。崔佩佩等[22]通过氮、磷和钾肥配施发现高粱根际微生物种类及微生物多样性有所增加。此外,小麦与苜蓿、豌豆、油菜间混作种植可促进小麦根际微生物数量增加,其中细菌数量最多,而真菌数量最少。
因此,土壤养分供给充足的条件下,由于扩散作用和根系吸收,使得根际土壤中的营养物质比非根际土壤浓度高,加之根系分泌物的释放诱导,有利于根区微生物繁殖,从而提高其丰富度和多样性,这也印证了本文NPK处理根际微生物多样性高,而养分缺乏处理根际微生物多样性低的结果。从已有相关的研究结果得知细菌多样性与土壤有机碳、有机氮含量正相关;而土壤真菌多为病原菌,不利于植物正常生长,可能会被植物根系主动过滤,导致根际真菌多样性比非根际低[23]。本研究结果显示,土壤pH、有效磷和速效钾含量对细菌群落结构的影响最为显著,而有机质和碱解氮含量对真菌群落结构的影响最为明显。对于根际与非根际土壤,李丽娟等[24]的研究表明,在植物根系的调控下,根际土中有机质与氮素、磷素以及钾素的相关性更为显著,而非根际土壤中仅钾素与碱解氮、有效磷呈显著负相关,其余各土壤养分因子之间均无显著相关性。本研究表明,与各处理非根际土壤养分含量相比,各处理根际土壤pH呈下降趋势,这是因为pH降低有利于土壤根际养分活化,从而出现有机质、有效磷及速效钾富集的趋势,而缓效钾之间差异不显著,这与背景值含量较高有关。
3.2 长期养分缺乏对冬油菜土壤微群落组成的影响
从本研究结果来看,长期氮磷钾养分供应不足导致冬油菜根际与非根际土壤在细菌、真菌群落组成上存在显著差异。由图1门水平分类结果来看,各处理根际土壤细菌的Patescibacteria相对丰度高于非根际土壤,而Rokubacteria的相对丰度则相反。真菌子囊菌门在-P、-K和NPK处理根际中的相对丰度均高于非根际土壤;担子菌门在-N1和NPK1土样中的相对丰度均较高。同时,在图2A属水平分类上,各处理非根际细菌的优势属种类比根际土壤显著增加,且非根际细菌的优势属相对丰度也高于根际土壤。非根际土壤的优势属,如Haliangium、Solibacter和Anaeromyxobacter在 根际土壤中的相对丰度显著降低,而Acidobacteria、Bryobacter和Gemmatimonas在根际土壤中富集并形成优势属,这与前人研究相一致[25],即根际养分差异会诱导优势菌群形成并与之匹配适应。具体而言,-N处理根际绿弯菌门相对丰度高于非根际土壤,这和Francioli等[26]的研究一致。变形菌门属富营养型细菌群落[27],因此,其在NPK肥处理根际的相对丰度高于非根际,这可能与根际营养充足有关。
在真菌门水平上(图1B),子囊菌门和担子菌门均为各处理的优势菌门,同时这两种真菌也是土壤有机质分解的重要参与者[28-29]。因此,养分缺乏土壤和NPK处理仍以担子菌门和子囊菌门为主导,这与前人研究结果相似[30-31]。尤其当土壤养分供应充足时,比较有利于富营养型菌群的生长;而当土壤养分贫瘠时,寡营养菌群相对丰度有增加的趋势[32-33],如本文-N和-P处理下根际壶菌门和球囊菌门的相对丰度显著高于NPK处理。
相关研究表明,根际与非根际菌群差异主要由pH和养分含量引起。通常条件下,根际微生物多样性高于非根际,一方面因为根际土壤有机碳、有机氮等养分含量高于非根际土,另一方面因为根际土壤可溶性盐含量低于非根际土,有利于特定类群增殖和提高丰度[34]。但长期肥料缺乏会加重根际营养不足,导致细菌和真菌优势菌群含量降低,以适应环境胁迫。此外,增施有机肥和秸秆还田、减少化肥农药施用量可显著改善土壤微生物种群的结构。有研究指出黄土旱地区作物连作时施适当化肥或有机肥,可提高土壤细菌多样性[35]。不同于小麦和水稻,冬油菜进入角果期,叶片逐渐脱落可增加土壤有机物质,能够为表层土壤微生物提供丰富的碳源;而长期肥料供应不足会导致冬油菜地上部生物量积累下降,进而影响土壤供碳能力和微生物繁殖。吴荣等[36]探究长期不同施肥方式对土壤微生物的影响,发现化肥与有机肥或者与秸秆配合施用的土壤细菌、放线菌数量比单施化肥的增幅效果明显。徐忠山等[37]的研究发现施用有机肥可不同程度增加细菌数量和种类多样性,进而使土壤系统向稳定健康的方向发展。因此,科学施肥有利于提高土壤微生物细菌和真菌的多样性,抑制优势菌群的形成,从而稳定土壤微生物生态系统的动态平衡。
综上所述,本研究结果表明,长期平衡施肥条件下,油菜根际土壤的细菌多样性比非根际土壤高,而真菌多样性低于非根际土壤,尤其长期不施用氮肥和磷肥会显著降低冬油菜根际细菌、真菌的丰富度。门分类水平上,油菜季土壤中的细菌优势物种是绿弯菌门、酸杆菌门和变形菌门;真菌优势物种是子囊菌门和担子菌门。相比NPK处理,-N、-P和-K处理对冬油菜根际、非根际细菌和根际真菌群落分布有显著影响,但对非根际真菌群落分布无明显作用。因此,长期养分缺乏会显著改变冬油菜根际细菌和真菌的群落组成结构及其多样性。
