长链脂酰辅酶A合成酶4调控环氧合酶2促进乳腺癌细胞转移的作用机制研究
2022-04-26林佳敏杨娜张苹苹徐邦牢
林佳敏 杨娜 张苹苹 徐邦牢
华南理工大学附属第二医院检验科(广州510180)
2020年我国新增乳腺癌确诊病例占全球24%,癌症相关死亡病例占全球30%,女性癌症患者乳腺癌的发病率已居首位[1]。研究表明,乳腺癌复发和转移相关的并发症是乳腺癌患者死亡的主要原因[2]。脂质代谢异常是癌细胞的重要标志之一,脂质代谢重编程为肿瘤转移提供了新的选择性优势[3-4]。游离脂肪酸在代谢之前必须经过酰基辅酶A 合成酶激活成酰基辅酶A 酯,长链脂酰辅酶A合成酶(ACSLs)家族的失调会改变细胞内游离脂肪酸的分布、种类和数量[5],从而导致癌症和其他代谢疾病,目前乳腺癌细胞转移过程中关键代谢酶的调控机制仍有待阐明。
长链脂酰辅酶A 合成酶4(ACSL4)是催化长链脂酸代谢的重要限速酶,属于ACSLs 家族中的一员。ACSL4 对长链多不饱和脂肪酸(polyunsaturated fatty acids,PUFA)如花生四烯酸(arachidonic acid,AA)的亲和力较高[6],AA 的代谢产物尤其是前列腺素E2(prostaglandin E2,PGE2)与多种生物过程有关[7]。环氧合酶2(cyclooxygenase-2,COX2)是AA 催化产生PGE2 重要的限速酶[8]。ACSL4 在侵袭性三阴乳腺癌表达更高,抑制ACSL4 可以减少细胞侵袭转移[9]。虽然有研究显示ACSL4 和COX2 对肿瘤增殖的影响[10],但其是否促进肿瘤的侵袭转移,仍有待进一步阐明。本研究旨在探讨ACSL4 是否通过调控COX2 促进乳腺癌转移,研究代谢关键酶ACSL4 调控脂酸代谢促进乳腺癌转移的作用,对开发乳腺癌转移的治疗新策略具有重要意义。
1 材料与方法
1.1 材料 人乳腺癌细胞MDA-MB-231、MCF-7 细胞株由本实验室保存;DMEM 高糖培养基(Gibco公司,美国)、胎牛血清(BI 公司,英国);新霉素和嘌呤霉素(索莱宝,中国)、0.25%胰蛋白酶(索莱宝,中国);Trizol(Invertrogen,美国);Transwell 小室(Corning 公司,美国);ACSL4 兔来源一抗(Abcam 公司,美国),COX2 兔来源一抗(CST,美国);VIMENTIN 兔来源一抗(CST,美国);GAPDH 鼠来源一抗(Proteintech,中国);PGE2 Elisa 检测试剂盒(Elabscience,中国);定量PCR 试剂(赛默飞,美国);逆转录酶(Takara,日本)。
1.2 实验方法
1.2.1 细胞培养 MDA-MB-231 和MCF-7 乳腺癌细胞,用含10%胎牛血清、1%双抗DMEM 高糖培养基,在37 ℃、含5%CO2的培养箱中培养细胞。
1.2.2 小干扰RAN(siRNA)转染和稳定细胞系建立 转染siRNA 由上海吉玛生物公司合成,siACSL4:sense5′-GAGGCUUCCUAUCUGAUUATT-3′,antisense5′ - UAAUCAGAUAGGAAGCCUCTT - 3′ ;MDA-MB-231 细胞转染按lipo-2000 的说明书进行。过表达ACSL4 慢病毒购于汉恒生物公司,按照公司说明书感染MCF-7 乳腺癌细胞,用1 mg/mL嘌呤霉素筛选。转染48 h,提取RNA 和蛋白质,分析基因水平和蛋白水平的变化。
1.2.3 RT-PCR 检测mRNA 表达 Trizol 法提取细胞的总RNA,调节RNA 浓度为1 000 ng/μL,按照Takara RNA 逆转试剂盒说明书进行逆转录,获得cDNA。采用SYBR Green PCR master mix 试剂盒进行qRT-PCR。根据2-ΔΔct值计算相对表达量。
1.2.4 Western blot 提取细胞总蛋白,加入5×loading buffer 后100 ℃水浴使蛋白变性,配置10%的SDS-PAGE分离胶和5%浓缩胶,分离蛋白(60 V,30 min 后换100 V,1 h),转膜(冰浴,100 V,1 h),封闭(5%脱脂奶粉,室温,1.5 h),使用一抗稀释液稀释ACSL4(1∶10 000)、COX2(1∶1 000)、VIMENTIN(1∶1 000)、GAPDH(1∶10 000)一抗,4 ℃过夜。二抗(1∶1 000)常温1.5 h,TBST洗膜后ECL显影。
1.2.5 ELISA PGE2 检测参照Elabscience 公司ELISA 试剂盒说明进行。
1.2.6 迁移实验 细胞生长到80%融合度时,用0.25%的胰酶消化,细胞消化后用无血清DMEM高糖培养基重悬细胞,调整细胞浓度为5 × 104个/mL。吸取200 μL 细胞悬液加入Transwell 小室上室,下室加入含10%血清的DMEM 完全培养基700 μL。24 h 后取出小室,去除培养基,4%多聚甲醛固定10 min 后,擦去小室上层的细胞,下层细胞用结晶紫染色30 min,在显微镜下拍照。
1.2.7 划痕实验 细胞生长到90%融合度时,用0.25%的胰酶消化,用完全培养基重悬细胞,铺板12 孔板,每个重复3 次,当24 h 细胞达到80% ~90%融合度时,使用10 μL 的枪头划痕,分别在0 h和24 h,显微镜下记录细胞迁移的面积。
1.3 统计学方法 用GraphPad 8.0 软件进行统计学分析,计量结果以()表示,计量资料两组对比采用独立样本t检验,组间比较采用单因素方差分析(one-way ANOVA),P<0.05 表示差异有统计学意义。
2 结果
2.1 ACSL4 对乳腺癌转移的影响 通过划痕实验和Transwell 实验发现,利用siRNA 瞬时敲低高侵袭性的MDA-MB-231 乳腺癌细胞ACSL4 表达后,其转移能力被显著抑制(P<0.05,图1A-B)。在低侵袭性的MCF-7 细胞中过表达ACSL4 后,其转移能力显著增强(P<0.05,图1C-D)。由此可见,内源性敲低或升高ACSL4 可以相应的降低或升高细胞的侵袭能力。
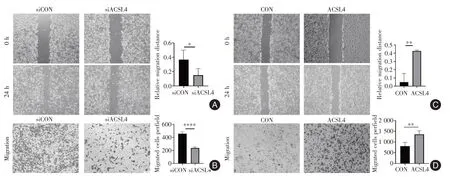
图1 ACSL4 对乳腺癌转移的影响Fig.1 Effect of ACSL4 on breast cancer metastasis
2.2 生物信息学分析 GEPIA 在线分析TCGA 数据库里,ACSL4 与COX2 表达呈正相关(r= 0.15,P<0.05,图2)。
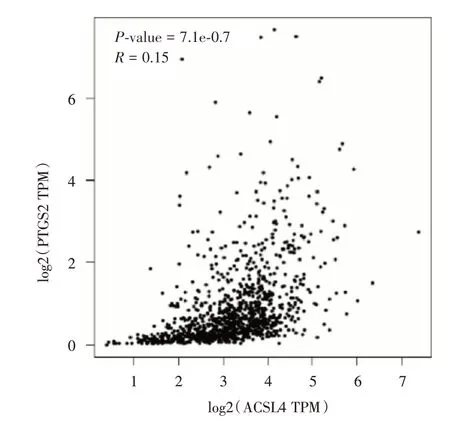
图2 TCGA 乳腺癌组织ACSL4 与COX2 基因表达相关性Fig.2 Correlation between ACSL4 and COX2 gene expression in TCGA breast cancer tissues
2.3 ACSL4 对乳腺癌细胞COX2 表达的影响 敲低高侵袭性的MDA-MB-231 乳腺癌细胞的ACSL4后,环氧合酶2(COX2)和波形蛋白(Vimentin)的基因表达水平和蛋白水平降低。过表达低侵袭性的MCF-7 细胞ACSL4 后,COX2 和波形蛋白表达水平升高(图3)。

图3 ACSL4 对乳腺癌细胞COX2 表达的影响Fig.3 Effect of ACSL4 on COX2 expression in breast cancer cells
2.4 ACSL4 对乳腺癌细胞PGE2 分泌水平的影响 敲低高侵袭性的MDA-MB-231 细胞的ACSL4后,通过ELISA 检测细胞分泌的PGE2 的含量,PGE2的分泌量下降了29.8%(图4A),在低侵袭性的MCF-7 乳腺癌细胞里过表达ACSL4,细胞分泌的PGE2 的含量升高29.5%,P<0.05(图4B)。
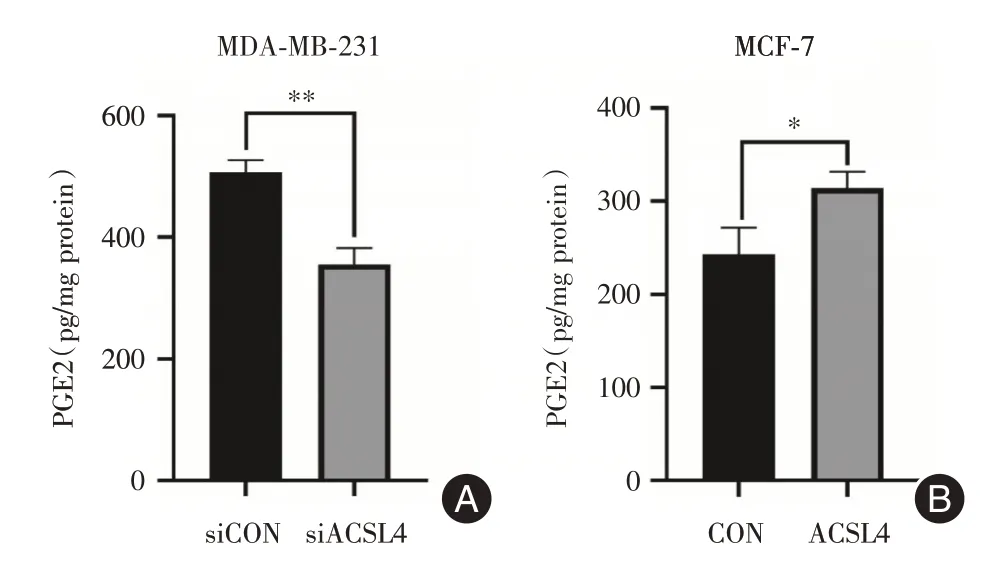
图4 ACSL4 对乳腺癌细胞PGE2 分泌水平的影响Fig.4 Effect of ACSL4 on PGE2 secretion level of breast cancer cells
3 讨论
ACSLs 家族是长链脂肪酸活化必需的酶(12 ~20 个碳原子),在哺乳动物中ACSLs 有五种亚型,即ACSL1、ACSL3、ACSL4、ACSL5 和ACSL6,它们在长链脂肪酸的激活中具有相似又特定的作用[11]。ACSL4 的偏好底物为二十碳多不饱和脂肪酸(PUFA),如花生四烯酸(AA)和二十碳五烯酸(EPA)[12],ACSL4 表达会增加游离AA 向PGE2 转化[13]。ACSL4 表达与乳腺癌细胞中的侵袭性表型相关,在侵袭性三阴乳腺癌中表达更多。17 β-雌二醇增加ACSL4 表达并促进AA 和EPA 的吸收。ASCL4 沉默阻止了17β-雌二醇诱导的p-Akt 和p-GSK3β 的上调和E-cadherin 的减少,从而减弱了癌细胞的侵袭能力[14]。虽然有研究表明,在乳腺癌侵袭性表型中,抑制ACSL4-LOX-COX2 通路可以显著降低肿瘤生长,但ACSL4 促进乳腺癌转移的具体机制还有待阐明[10]。本研究显示ACSL4能明显增强乳腺癌细胞系的迁移和侵袭能力。内源性ACSL4 的缺失或升高,可以分别抑制或提高MDA-MB-231 和MCF-7 细胞的侵袭能力,提示ACSL4 是乳腺癌细胞侵袭促进因子,为乳腺癌的治疗提供了新的治疗靶标。
AA 通过三种不同酶催化的途径产生不同的生物活性产物:环氧合酶(COX)、脂肪氧化酶(LOX)和细胞色素P450(CYP)通路[15]。COX 家族成员,包括COX1 和COX2,介导AA 转化为PGH2,PGH2 随后代谢为PGD2、PGE2、PGF2α、PGI2,和血栓素A2(TXA2)[16]。源自COX2 的PGE2 可以激活结肠癌中雷帕霉素复合物1(mechanistic target of rapamycin complex 1,mTORC1)信号转导的机制靶点,从而促进VEGF 的表达和释放,引起细胞增殖能力增强和血管生成[17]。另一项研究表明,PGE2激活EP1R,从而通过涉及蛋白激酶C(PKC)、NF-κB和叉头盒蛋白C2(FoxC2)的信号传导增加β1-整联蛋白的表达,诱导癌细胞转移[18]。本研究通过生物信息学分析了ACSL4 与COX2 存在正相关关系,在MDA-MB-231 细胞中敲低ACSL4,可以在mRNA 水平和蛋白水平上降低COX2 的表达,同时,通过ELISA 检测细胞分泌的PGE2 的水平减低,在MCF-7 细胞中过表达ACSL4,可以在蛋白水平上提高COX2,通过ELISA 检测细胞分泌的PGE2的水平升高,与生物信息学预测结果一致,进一步提示ACSL4 通过调控COX2 的表达,催化AA 生成PGE2,增加PGE2 的生成。
波形蛋白是间充质细胞中的一种Ⅲ型中间丝蛋白,主要在成纤维细胞、内皮细胞和淋巴细胞中表达[19],在细胞水平上,波形蛋白参与细胞迁移、分化、增殖、粘附和侵袭[20]。有研究表明,波形蛋白高表达与乳腺癌淋巴结转移有一定的相关性,是影响乳腺癌预后的潜在因素[21]。本研究结果显示在MDA-MB-231 细胞中敲低ACSL4,可以在mRNA 和蛋白水平降低波形蛋白的表达,在MCF-7细胞中过表达ACSL4,可以在蛋白水平上升高波形蛋白的表达。进一步提示了ACSL4 与乳腺癌转移密切相关,ACSL4 可能是通过调节COX2 促进PGE2 的分泌,增强了乳腺癌细胞的侵袭转移的能力,这可能是乳腺癌侵袭转移的机制之一。
综上所述,本研究目前在细胞水平揭示了ACSL4 可以促进乳腺癌细胞的迁移和侵袭能力,其主要通过调节COX2 促进PGE2 分泌发挥作用。下一步需要在动物体内实验验证ACSL4-COX2 在乳腺癌转移过程中的作用,深入研究ACSL4 调节COX2 的分子机制以及代谢产物PGE2 涉及的信号通路对乳腺癌发生发展的影响,以期开发针对乳腺癌转移的治疗新策略。
