MicroRNA-99b-5p通过抑制成纤维细胞生长因子21表达促进心肌细胞肥大
2022-04-18陈丽文陈泽润朱杰宁徐金东郭惠明单志新
陈丽文,郭 晶,陈泽润,朱杰宁,徐金东,郭惠明,单志新,,,王 晟
(1.广东省心血管病研究所//广东省人民医院//广东省医学科学院,广东广州 510080;2.华南理工大学医学院,广东广州 510006;3.南方医科大学第二临床医学院,广东广州 510280)
近年来,我国心血管疾病引发的死亡率和发病率仍呈增长趋势[1]。心肌肥厚是多种心血管疾病发生的病理基础,主要表现为心肌细胞在长度和宽度上的改变,即心肌细胞肥大,而非数量上的增加[2]。心肌细胞发生病理性肥大时会出现心脏胎儿基因的重新激活和表达,如心房钠尿肽(atrial natriuretic peptide,ANP)以及β 肌球蛋白重链(β myosin heavy chain,β-MHC)等的编码基因[3]。在心肌肥厚的早期阶段,心肌收缩力增加是一种短期的适应机制,而持续的刺激将导致适应性不良的心脏重构,并最终发展为心力衰竭[4]。目前,心肌肥厚及后续转变为心力衰竭的确切分子机制尚未明确,有待进一步的研究证实。越来越多的研究表明微小RNA(microRNA,miRNA)参与调节心肌细胞的肥大[5]。MicroRNA 通过与靶基因mRNA 结合,抑制靶基因的翻译,进而发挥调节相关生物学过程的重要作用[6]。多种microRNA 可靶向调节与心脏疾病相关的基因表达[6-7],如心肌特异性敲除miR-22 可减轻应激诱导的心肌肥厚[8];miR-206 可被YAP 上调,并介导YAP 诱导的心肌细胞肥大[9]。我们以往的研究证实miR-199a-3p 和miR-199b-5p 可分别通过结合Rb-1 和CDK9,发挥促进心肌细胞肥大的作用[10-11]。成纤维细胞生长因子21(fibroblast growth factor 21,Fgf21)是一种可在多种的组织器官中表达并发挥多种起生物学功能的分泌蛋白[12]。有研究显示在异丙肾上腺素诱导肥厚的小鼠心肌,增加Fgf21 表达能够减轻小鼠心肌肥厚的发生,其可能机制是Fgf21 通过增加SIRT1 表达来减弱血管紧张素Ⅱ(angiotensin Ⅱ,Ang Ⅱ)诱导的心肌肥厚[13],但Fgf21 发挥心脏保护作用的分子机制尚不完全清楚。我们的前期工作已证实circRNA_005647 可作为分子海绵吸附miR-99b-5p来抑制心肌细胞发生肥大[14],但miR-99b-5p在心肌肥大中的具体作用机制尚不明确。本文旨在探讨miR-99b-5p 对小鼠心肌细胞(neonatal mouse ventricular cardiomyocytes,NMVCs)肥大相关基因表达的促进作用,并阐明Fgf21 是否介导miR-99b-5p 发挥对心肌肥大表型的调节作用。
1 材料与方法
1.1 组织标本
利用RT-qPCR 检测并比较心衰(heart failure,HF)患者、健康器官捐献者心肌组织标本中miR-99b-5p 的表达水平。临床患者组织样本由广东省心血管病研究所提供,开展的实验均经过广东省人民医院伦理委员会审批[批准号No.GDREC201923 8H(R1)],心衰及健康对照患者病例资料信息同以往报道[15]。
1.2 实验动物
C57BL/6 乳小鼠(出生日龄1~3 d,雌雄不限)和8周的性成熟雄性C57BL/6小鼠由广州中医药大学实验动物中心提供,属于无特定病原体级别动物。开展的动物实验通过广东省人民医院伦理委员会审批[批准号:粤医科伦理2017015A 号],实验动物许可证号为SCXK(粤)2013-0034。
1.3 主要试剂
TRizol 总RNA 提取裂解液(Invitrogen);逆转录试剂(TaKaRa);miR-99b-5p、U6、Fgf21等序列引物(Invitrogen);2× SYBR Green Pro Taq HS Premix(艾科瑞);DMEM/F-12 无血清培养基、0.25%EDTA 胰蛋白酶(Gibco)、特级澳洲胎牛血清;细胞转染试剂Lipofectamine 2000、Oligofectamine(Invitrogen);miR-99b-5p mimic、Fgf21 siRNA、超氧化物歧化酶2(Superoxide dismutase,Sod2)siRNA(广州锐博);RIPA蛋白裂解缓冲液(碧云天);SDS-PAGE凝胶配制试剂盒(碧云天);4×loading buffer、蛋白marker 26616(Thermo Fisher Scientific);BCA 蛋 白浓度测定试剂盒;PVDF 膜(Whatman);抗心房钠尿肽抗体(Bioworld);抗β 肌球蛋白重链抗体(Sigma);抗FGF21 抗体(SAB 公司);抗SOD2 抗体(Cell Signaling Technology)、抗GAPDH 抗体(Protein Technology);萤光素酶活性检测试剂盒(Promega);抗氧化 剂N-Acetyl-L-cysteine(MCE 公司)等。
1.4 主要方法
1.4.1 原代乳小鼠心肌细胞的分离与培养 取SPF级C57BL/6乳小鼠心脏于高压灭菌后的PBS溶液中漂洗去除残血,并修剪去除血管等其他组织,转移至干净的离心管中,加入0.25% EDTA 胰蛋白酶及PBS 溶液,于4 ℃摇床10 h。次日于装有乳小鼠心脏的离心管加入完全培养基,放入37 ℃中温浴10 min 停止消化。巴氏管将心脏吸至新的离心管,加入不含血清的细胞培养基及胶原酶Ⅱ,37 ℃中轻摇10 min。待组织吹散后加入完全培养基进行离心,用干净PBS 溶液重悬下面一层的细胞后,经滤网(70 μm 孔径)过滤至新的离心管里后进行离心,重旋转至培养瓶。1.5 h 后将瓶中的悬液吸出,分至细胞培养板中培养。
1.4.2 C57BL/6 小鼠横向主动脉弓缩窄模型建立 利用横向主动脉弓缩窄(transverse aortic restriction,TAC)法建立小鼠心肌肥厚模型。将小鼠置于诱导麻醉盒中,2%异氟烷吸入诱导麻醉,麻醉后仰卧位固定,以第二肋骨为中心剪开表皮,从第二肋骨与胸骨连接处剪开暴露主动脉弓,于无名动脉和左颈总动脉结扎主动脉弓,待小鼠情况稳定后缝合,术后进行连续3周的观察和饲养。
1.4.3 原代乳小鼠心肌细胞转染miR-99b-5p 和siRNA 将NMVCs 于12 孔细胞培养板稳定培养后,在EP管中加入Oligofectamine(1.75 μL/孔)与无血清培养基混合,在另外的EP 管中加入mimic 或siRNA(100 nmol/L)与无血清培养基混合,将Oligo与mimic 或siRNA 轻轻混匀,静置包裹15 min。将NMVCs 用PBS 缓冲液清洗后更换为含3 %血清的培养基,再加入包裹好的混合物,置于体积分数5%CO2、37 ℃培养箱培养36 h。
1.4.4 Fgf21和Sod2重组腺病毒的构建和包装 参照已报道方法[16],分别将小鼠Fgf21 和Sod2 基因编码区序列定向插入pAd-Track-cmv 载体,并继续进行后续的相应重组腺病毒质粒载体构建和重组腺病毒的包装及扩增。实验时以rAd-GFP 作为对照组腺病毒(rAd-Fgf21、rAd-Sod2 和rAd-GFP 的MOI均是5)。
1.4.5 麦胚凝集素染色 麦胚凝集素(triticum vulgaris lectin,WGA)染色参照我们已报道的方法[15],并略有改进。
1.4.6 实时荧光定量PCR TRIzol 法提取NMVCs或小鼠心肌组织总RNA,20 μL mRNA 逆转录体系中加入5×Prime Script RT Master Mix 4 μL 以及1.0 μg 总RNA 模板进行逆转录,获得相应的cDNA,以GAPDH 为内参照检测相关基因mRNA 表达水平。MiRNA 成熟体逆转录体系中加入5×PrimeScript RT Buffer、RT Enzyme Mix、miR-99b-5p RT 引物、U6 RT 引物,以1.0 μg总RNA为模板进行成熟体逆转录,获得相应产物用于检测miR-99b-5p 表达水平。利用ViiA 7 Quantitative PCR System(Applied Biosystems)进行实时荧光定量PCR 并分析得到的实验结果。计算相应的2-ΔΔCt值,比较不同实验分组间miR-99b-5p、Fgf21 以及肥大相关基因的表达水平。相应的引物序列列于表1。

表1 PCR引物序列Table 1 The sequences of the primers for PCR
1.4.7 蛋白质免疫印迹法 利用含蛋白酶抑制剂的RIPA 裂解缓冲液裂解细胞培养板中的NMVCs或小鼠心肌组织(组织需进行研磨)后,高速离心,测蛋白浓度。等质量分装蛋白后再与4×SDS loading buffer震荡混匀,放入99 ℃进行变性10 min。按相应的分组,将样品加入凝胶梳孔中,电泳分出大小差异的蛋白。取出凝胶后,在电转缓冲液中将蛋白转印至PVDF 膜上。封闭后利用相应的一抗[β-MHC(1:1 000)、ANP(1:1 000)、FGF21(1:2 000)、SOD2(1:1 000)、GAPDH(1:5 000)]孵育4 ℃过夜,TBST 液清洗后敷上相应的二抗,1 h 后TBST再次清洗,即可进行化学发光反应。
1.4.8 双萤光素酶报告基因检测 将Fgf21 的3’UTR 序列及其与miR-99b-5p 潜在的结合位点突变后的3’UTR序列分别构建入pGL3-promoter载体中。在构建好重组质粒后,将相应质粒pGL3-promoter-Fgf21、pGL3-promoter-Fgf21-mut 以及miR-99b-5p mimic 与内参照pRL-TK 质粒共转染至HEK293 细胞中,24 h 后测定萤火虫萤光素酶(FL)和海肾萤光素酶(RL)的活性。通过计算比较FL/RL比值的大小,判断miR-99b-5p 与Fgf21 的结合作用。
1.4.9 鬼笔环肽染色 将原代乳小鼠心肌细胞于Confocal皿中培养,进行实验干预后,用干净PBS清洗。在血中加入40 g/L 多聚甲醛溶液进行固定。然后更换成PBS溶液,置于摇床上轻轻漂洗。接下来,配制0.1% TritonX-100 透化液,加入皿中透化处理细胞,再次漂洗。加入鬼笔环肽工作液后避光,细胞染色60 min。染色完成用PBS 洗后,加入含DAPI 封片剂,在4 ℃冰箱用锡箔纸避光保存,待激光共聚焦显微镜观察心肌细胞形态。
1.5 统计学方法
图像处理软件Image J 分析蛋白条带,利用GraphPad Prism8、SPSS 21.0 进行统计分析。计量资料结果用均数±标准差表示。两组间比较采用t检验;方差不齐的两组数据比较用Welch’st检验;多组间比较采用单因素方差分析;采用Bonferroni校正的t检验进行组间两两比较。P<0.05时,差异具有统计学意义。
2 结果
2.1 MiR-99b-5p在发生肥厚的心肌中表达增加
与对健康捐献者心肌相比,WGA 染色显示心衰患者心肌细胞的横截面积明显增大(P<0.001;图1A),同时心衰患者心肌中miR-99b-5p 表达水平显著升高(P<0.05;图1B)。我们将C57BL/6 小鼠分为假手术组(Sham)和TAC 手术组,并进行相应的手术处理。WGA 染色显示TAC 手术小鼠心肌细胞的横截面积明显增大(P<0.01;图1C),并且TAC手术小鼠心肌中miR-99b-5p水平显著增加(P<0.01;图1D)。我们用AngⅡ进行处理NMVCs,建立心肌细胞肥大模型。鬼笔环肽染色显示,AngⅡ处理的NMVCs 细胞明显增大(P<0.05;图1E),RT-qPCR 结果显示发生肥大的NMVCs 中miR-99b-5p水平显著上调(P<0.01;图1F)。

图1 miR-99b-5p在肥厚心肌中表达增加Fig.1 Upregulation of miR-99b-5p in the hypertrophic myocardium
2.2 MiR-99b-5p 促进原代乳小鼠心肌细胞中肥大相关基因表达
鬼笔环肽染色结果显示转染miR-99b-5p mimic 的NMVCs 细胞面积明显增大(P<0.05;图2A)。RT-qPCR 和Western blot 检测发现,转染miR-99b-5p mimic的NMVCs中肥大相关基因Myh7和Anp 表达显著增加(P<0.01 或P<0.001;图2B、2C)。
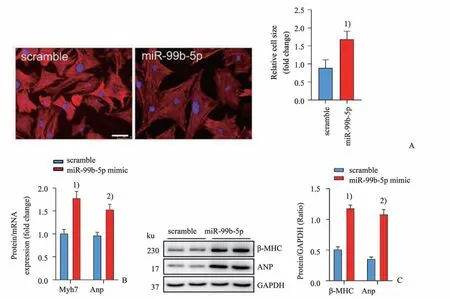
图2 miR-99b-5p促进NMVCs中心肌肥大相关基因表达Fig.2 MiR-99b-5p enhances hypertrophy-related gene expression in NMVCs
2.3 MiR-99b-5p靶向Fgf21发挥促进NMVCs中心肌肥大相关基因表达的作用
生物信 息学分 析(https://www.mirbase.org/;https://starbase.sysu.edu.cn/)结果提示Fgf21 基因的3’UTR 存在与miR-99b-5p 潜在结合的位点(图3A)。进一步的双萤光素酶报告基因实验证实miR-99b-5p与Fgf21 3’UTR间结合作用,经单因素方差分析,四组间差异有统计学意义(F=4.976,P=0.011 7),采用Bonferroni 法进一步作两两比较,发现miR-99b-5p 与Fgf21 3’UTR 间存在特异结合作用(P<0.01),而突变3’UTR 的相应结合序列后,miR-99b-5p 则不能降低萤光素酶的活性(图3B)。RT-qPCR 结果显示,FGF21 mRNA 在心衰病人心肌中表达降低,但差异无统计学意义(图3C),而Fgf21 mRNA 在TAC 手术诱导肥厚的小鼠心肌中表达显著降低(P<0.05;图3D)。转染miR-99b-5p mimic 可在RNA 和蛋白水平抑制NMVCs 中Fgf21表达(P<0.01;图3E,3F)。
Western blot 结果显示,Fgf21 siRNA 和miR-99b-5p mimic 可一致性地增强NMVCs 中β-MHC和ANP蛋白表达(P<0.000 1;图3G)。而利用腺病毒介导在NMVCs 过表达Fgf21 可显著抑制NMVCs中肥大相关基因Myh7 和Anp 表达(P<0.001),并逆转miR-99b-5p对心肌细胞肥大相关基因表达的促进作用(P<0.0001;图3H)。

图3 miR-99b-5p靶向Fgf21发挥促进NMVCs中心肌肥大相关基因表达作用Fig.3 Fgf21 mediates the pro-hypertrophic effect of miR-99b-5p on NMVCs
2.4 Fgf21/Sod2 轴介导miR-99b-5p促进NMVCs中心肌肥大相关基因表达作用
Western blot 结果显示,ROS 抑制剂NAC 与Fgf21 能一致地有效逆转miR-99b-5p 促心肌细胞肥大的作用(P<0.01 或P<0.001;图4A)。利用腺病毒介导过表达Fgf21 和Sod2 均能显著抑制NMVCs 中β-MHC 和ANP 表达(P<0.01),并且Fgf21 可增加NMVCs 中SOD2 表达(P<0.05 或P<0.000 1;图4B)。用siRNA 敲减Fgf21 的NMVCs 中SOD2 表达显著降低(P<0.000 1),而沉默Fgf21 和Sod2 表达则可与miR-99b-5p mimic 一致地促进NMVCs 中β-MHC 和ANP 的表达(P<0.05 或P<0.01;图4C)。
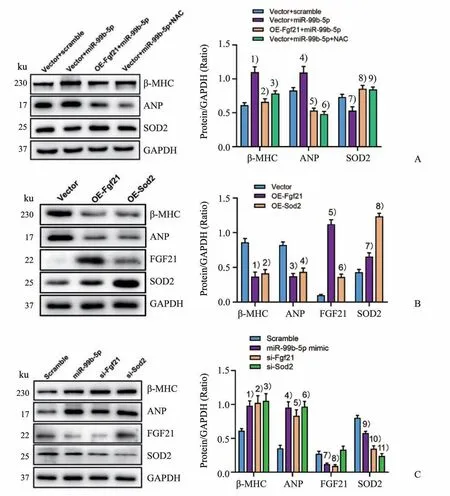
图4 Fgf21/Sod2轴介导miR-99b-5p促进NMVCs中心肌肥大相关基因表达作用Fig.4 Fgf21/Sod2 axis mediates the pro-hypertrophic effect of miR-99b-5p on NMVCs
3 讨论
在本文中,我们证实miR-99b-5p 在心力衰竭患者心肌、TAC 诱导的小鼠心肌以及AngⅡ诱导的心肌肥厚细胞模型中均表达增强,提示miR-99b-5p 可能参与心肌肥厚的过程。功能实验的结果证实转染miR-99b-5p可特异增加小鼠心肌细胞面积和心肌肥大相关基因表达,具有促进心肌细胞肥大的作用。本研究明确了介导miR-99b-5p发挥促心肌细胞肥大作用的下游靶基因,为阐明circRNA_005647 通过结合miR-99b-5p 来抑制心肌细胞肥大的作用机制提供科学依据。
已有研究显示miR-99b-5p参与人成骨细胞增殖、骨关节炎以及非小细胞肺癌转移等过程[17-19],但尚未见miR-99b-5p参与心肌细胞相关表型调控的报道。我们通过miRNA 数据网站的TargetScan、miRanda 和PicTar 等软件共同预测到Fgf21 是miR-99b-5p 的潜在靶基因,鉴于已有报道显示Fgf21 可以调控miR-143[20]和miR-27b[21]发挥调节心血管疾病进程的作用,但尚未见有关miRNA 调控Fgf21表达的报道,因而本文拟探讨miR-99b-5p 是否可通过调节Fgf21 表达发挥促进心肌细胞肥大作用。本文通过双萤光素酶报告基因实验显示miR-99b-5p 可 与Fgf21 基因的3’UTR 特异结 合,并明确miR-99b-5p 可在转录水平抑制Fgf21 表达。基于心衰病人心肌和TAC 小鼠心肌中miR-99b-5p 和Fgf21 mRNA 表达的检测结果也提示两者在心肌肥厚过程中表达负相关。功能实验证实在细胞水平敲减Fgf21 可与miR-99b-5p 一致性地抑制心肌细胞肥大表型,而过表达Fgf21 可有效逆转miR-99b-5p的促心肌细胞肥大作用,因此Fgf21介导了miR-99b-5p 的促进心肌细胞肥大作用。本文上述结果与以往Fgf21 具有抑制心肌细胞肥大作用的报道相符[13]。
长时间的病理性刺激可加重心肌的氧化损伤,而氧自由基清除剂和抗氧化剂可以减轻氧化应激所致的亚细胞功能缺损和心脏功能的降低[22]。已有研究证实Fgf21 可减轻心肌受到的氧化损伤作用[23],因此本文试图明确miR-99b-5p 是否通过靶向Fgf21 来调节心肌细胞的氧化应激和肥大表型。我们发现抗氧化剂NAC 和过表达FGF21 均能一致抑制miR-99b-5p 的促心肌细胞肥大的作用,提示miR-99b-5p 通过增加氧化应激途径来促进心肌细胞肥大。既往研究表明,SOD2 是细胞线粒体内中和超氧阴离子自由基(·O)的主要抗氧化酶,在维持正常心脏功能和抑制心肌肥厚表型中发挥重要作用[24-26]。已有报道Fgf21 可在mRNA 水平上促进抗氧化基因Sod2 表达[21],而本文进一步在蛋白水平证实Fgf21可调控NMVCs中Sod2表达,并从功能上明确Fgf21/Sod2 轴介导了miR-99b-5p 的促心肌细胞肥大作用。
综上,本文证实了miR-99b-5p 通过抑制其下游靶基因Fgf21,进而抑制其下游Sod2 基因表达来促进心肌细胞肥大。在后续的研究中,我们将继续探讨Fgf21/Sod2 轴介导miR-99b-5p 促心肌细胞肥大的分子机制,并在整体水平上明确Fgf21 介导miR-99b-5p 发挥促进心肌肥厚的作用机制。
