弥可保预防长春新碱诱导的病理性疼痛的机制
2022-04-18黄睿臻魏绪红
黄睿臻,徐 劲,魏绪红
(中山大学中山医学院生理教研室//疼痛研究中心,广东广州 510080)
长春新碱和铂类药物等癌症化疗药物的使用常常会引起外周神经病变(chemotherapy-induced peripheral neuropathy,CIPN)。CIPN 主要表现为本来不引起疼痛的机械刺激及冷热刺激所诱发的疼痛[1]。CIPN 限制化疗药物的使用并严重影响病人的生活质量[2],目前尚没有较好的治疗方法。因此,我们需要另一种新的有效的治疗或预防CIPN的策略。病人腓肠神经活检和动物实验发现,化疗导致外周传导触压觉的有髄传入纤维丢失[3],这可以解释化疗病人触压觉迟钝。但是,肢体末梢麻木伴有剧烈疼痛的机制则不清楚。众所周知,慢性疼痛是由C 纤维传导的,而C 纤维又分为肽能和非肽能两大类,前者表达IB4,投射到脊髓背角Ⅱ板层的内侧,而后者表达降钙素基因相关肽(calcitonin gene related peptide,CGRP)、神经生长因子受体TrkA和P物质,投射到脊髓背角Ⅰ板层和Ⅱ板层的外侧,直接与背角表达神经激肽-1 受体(neurokinin-1 receptor,NK-1R)的神经元形成突触[4]。NK-1R 阳性神经元投射到高位中枢(丘脑和臂旁核)。研究表明,选择性地去除脊髓背角NK-1R 阳性投射神经元可防止炎性痛和神经病理性疼痛[5]。我们之前的研究表明,长春新碱可引起脊髓背角CGRP 非肽能C 纤维芽生,但不能引起IB4肽能C 纤维芽生,这一变化可能是引起化疗后疼痛产生的重要原因[6]。弥可保(甲钴胺,McB)是包含钴元素的一种维生素B12,对神经组织有很强的亲和作用。弥可保对神经系统有着重要的功能,如,促进神经修复[7]。弥可保也可以显著提高DNA 甲基化,并已有报道其可以调节糖尿病神经病理性疼痛[8]、神经损伤[9]、慢性压迫性神经损伤所引起的痛觉过敏[10]或带状疱疹后遗神经痛[11]。目前弥可保用于治疗糖尿病引起的周围神经病变[12]。但另一项研究报道,弥可保对腰椎狭窄引起的疼痛无效[13]。我们之前的研究报道,弥可保能防止长春新碱诱导的神经病理性疼痛,但其机制目前仍不清楚。脊髓背角C 纤维诱发电位被认为是神经病理性疼痛的突触模型,在本部分内容中,我们将研究预防性给予弥可保对长春新碱诱导的脊髓背角C 纤维诱发电位的影响,并进一步观察弥可保对脊髓背角核因子κB(NF-κB)信号通路激活的影响。
1 材料与方法
1.1 实验动物
实验采用成年Sprague-Dawley(SD)大鼠,雄性,180~200 g,购于中山大学实验动物中心,动物的使用协议和动物处理程序获得了中山大学实验动物管理与使用委员会(IACUC)的批准。动物被分笼饲养并能够自由获取标准啮齿动物食物和水,室温保持在(25 ± 1)℃,湿度保持在50%~60%,并处于12 h白天-黑夜循环照明的环境。所有的动物被随机分配到不同的处理组。
1.2 药品
长春新碱(深圳万乐药业有限公司,中国)用无菌生理盐水配成50 μg/mL,密封、遮光保存于4 ℃;以每天0.1 mg/kg 的剂量对大鼠腹腔注射上述长春新碱溶液,连续给药10 d,每天1次[6]以建立化疗药物诱导的疼痛模型。弥可保注射液(0.5 mg/mL,卫材株式会社,日本)密封、遮光保存于4 ℃。因化疗性药物诱导的疼痛治疗难度大于预防性给药,我们选择于腹腔注射长春新碱造模前的15 d开始预先使用上述弥可保注射液对大鼠进行预防性给药,以1 mg/kg[14]的剂量每隔一天腹腔注射给药1次,直到最后1 次长春新碱给药后的第9 d(即第1次给长春新碱后19 d),如两个药物需要同一天给药,则弥可保在长春新碱注射之前30 min给药。具体给药方式见图1。
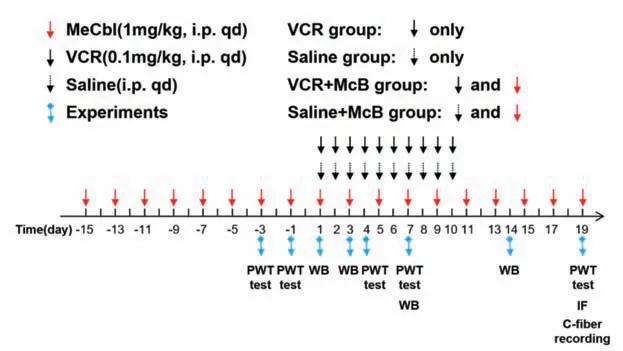
图1 大鼠实验分组给药及其实验情况Fig.1 The drug administration in different groups of rats and the experimental timeline
1.3 50%机械刺激撤足阈值测定
于测试前3 d将大鼠放置于透明有机玻璃箱内适应环境,每次20 min 以上,每天1 次。正式测试时待大鼠安静后,采用Chaplan 等[15]提出的Up-Down 方法,选10 根强度呈对数递增方式的纤维毛(von Frey hairs,3.84,4.08,4.17,4.31,4.56,4.74,4.93,5.07,5.18,5.46,Aesthesio,Italy)分别对大鼠后肢左右脚足心部进行刺激,每次刺激持续时间为6~8 s。大鼠出现快速撤足和(或)舔足反应则为撤足反应阳性,如无出现则为撤足反应阴性。以对数值3.84(压力0.692 g)刺激强度为初始刺激强度,若撤足反应为阴性则选用相邻递增的刺激强度的纤维毛继续刺激,若撤足反应为阳性则选择相邻递减的刺激强度给予刺激。50%机械刺激撤足阈值的计算算式:50%机械刺激撤足阈值(50% Paw withdrawal threshold,式中Xf为最末次测试von Frey hair的对数值;k值可根据撤足反应模式查表(Chaplan 等[15]提出的Up-Down 方法)得出;δ为10 根von Frey hair 间对数差值的均值。
1.4 电生理记录和坐骨神经刺激
在第19 d 分别取生理盐水组、长春新碱组、长春新碱+弥可保组、生理盐水+弥可保组的大鼠,每组5~7 只用乌拉坦(1.5 g/kg)腹腔注射麻醉;在腰4~6 间行椎板切除术,暴露脊髓腰膨大部位。游离左侧坐骨神经,使用双极氯化银电极刺激坐骨神经。除用于记录的脊髓节段外,暴露的神经组织均用37 ℃石蜡油覆盖。应用本实验室常规方法记录脊髓背角C 纤维诱发电位,记录方法如文献所述[16]。通过氯化银电极电刺激游离的坐骨神经以诱发脊髓背角场电位。玻璃微电极(电阻0.5~1 MΩ)进行细胞外记录,用电控的微量推进器(Narishige scientific instrument laboratory)将电极深度控制在脊髓表面以下300~500 μm。用数/模转换器(ADC-42.PICO)以10 kHz 的速度处理和储存数据。以C 纤维诱发电位的幅度作为参数。单方波(1~20 V,0.5 ms)刺激分离的左侧坐骨神经诱发脊髓背角场电位,记录曲线稳定后静置15 min,随后分别以1 V、2.5 V、5 V、7.5 V、10 V、12.5 V、15 V、20 V、26 V 的刺激强度,每个刺激强度刺激坐骨神经3次,每次刺激间隔1 min,以记录不同刺激强度对C纤维诱发场电位幅度的影响,坐骨神经的刺激点到脊髓背角的记录点距离约为10 cm。
1.5 免疫荧光实验
分别取生理盐水组、长春新碱造模组、长春新碱+弥可保组、生理盐水+弥可保组的大鼠,经腹腔注射乌拉坦(1.5 g/kg)麻醉后,剪开胸腔,灌注针经心尖垂直插入左心室至主动脉,快速灌注生理盐水300 mL,再用40 g/L 的多聚甲醛300 mL 灌注,解剖大鼠取出脊髓腰膨大段,再放入40g/L 的多聚甲醛中后固定1 h,随后转入30%蔗糖中脱水3 d。标本经蔗糖脱水后进行冰冻切片(LEICA CM1900),脊髓切片厚度为25 μm,4 ℃短暂保存。收集脊髓冰冻切片后用0.01mol/L PBS 洗3 次,每次10 min,然后室温下用免疫染色封闭液(Beyotime Biotechnology,China)作用于切片1 h。随后加入对应的一抗,分别为anti-calcitonin gene related peptide,anti-CGRP(1:200;rabbit;CST,USA),anti-phosphorylated NF-κB p65(Ser311;1:200;rabbit;Abcam,UK),anti-IL-10(1:100;rabbit;Abcam,UK),anti-NeuN(1:200;mouse;Abcam,UK),anti-GFAP(1:600;rabbit;Abcam,UK),anti-Iba-1(1:500;rabbit;Abcam,UK),摇床4℃孵育12 h,洗去一抗,用0.01M PBS 漂洗,每次10 min,共3 次,加入与一抗对应的二抗,分别为Cy3-conjugated(1:400;Jackson ImmunoResearch,USA),Alexa 488-conjugated(1:500;Jackson ImmunoResearch,USA),避光室温下慢摇1 h,随后洗去二抗,再用0.01mol/L PBS 洗4次,每次10 min,随机挑选切片贴于防黏附载玻片上,滴上含防荧光猝灭剂的封片液(Southern Biotech,USA)并用盖玻片轻轻封片,防止气泡产生。将制作好的玻片于4 ℃下避光晾干后立即于荧光显微镜(Leica DFC350 FX camera,Heidelberg,Germany)下观察并拍照。组织切片免疫荧光染色结果的半定量分析使用Image Pro 软件,计算每张切片上免疫阳性区域而获得。随机挑选5~6张切片的拍照结果,导入Image Pro 软件分析阳性区域大小,用阳性区域的面积比上脊髓背角的总面积;以及计数脊髓背角神经元、星形胶质细胞、小胶质细胞的数量,分别用p-p65 阳性细胞数比上神经元、星形胶质细胞、小胶质细胞的数目。
1.6 Western blot实验
在第1、3、7、14 d 时,分别取生理盐水组、长春新碱组的大鼠,每组5~7只用乌拉坦(1.5 g/kg)腹腔注射麻醉动物麻醉后,解剖大鼠并快速取出腰膨大段脊髓置于液氮中冷冻,然后加入SDS裂解液(Sigma-Aldrich,USA)(10 μL/mg)和蛋白磷酸酶抑制剂(Roche,Switzerland)匀浆并超声破碎,13 600×g4 ℃离心后取上清液,上清用SDS-PAGE进行分离,并转至PVDF膜(Sigma-Aldrich,USA)上。1%BSA室温(Sigma-Aldrich,USA)封闭1 h,然后再在4℃环境下 与一抗(phosphorylated NF-κB p65;Ser311;rabbit;1:1 000;Abcam;UK or NF-κB p65;rabbit;1:1 000;Abcam;UK or β-actin;rabbit;1:1 000;Abcam;UK)一起轻摇孵育过夜。第2 天用TBST 洗4 遍,随后用辣根过氧化物酶标记的二抗(goat anti-rabbit IgG,1:10 000;Abcam,UK)孵育1 h,TBST洗4遍,经ECL发光液(Shanghai Epizyme Biomedical Technology,China)显色曝光,再经化学发光成像系统(Tanon,China)内置CCD 摄像头拍摄显影条带,使用Image-Pro Plus 软件对图像进行灰度分析。对每个目的条带进行总灰度值的分析,并用β-actin的总灰度值进行标准化。
1.7 统计学分析
通过graphpad 6.0 软件进行统计学分析,实验结果以均数±标准误(means ± SEM)表示。首先用Shapiro-Wilk 检验数据是否符合正态分布,P>0.05被认为符合正态分布。二组计量资料的均数比较,如果每一组资料都呈正态分布并且方差齐性,组间比较采用t检验,反之用秩和检验;不符合正态分布的数据采用非参数检验。多组数据组间比较采用单因素方差分析(one-way ANOVA)继以Tukey 检验。P<0.05被认为有统计学意义。
2 结果
2.1 连续腹腔注射长春新碱引起机械痛敏
与手术前基础值比较,腹腔注射长春新碱(vincristine,VCR,0.1mg·kg-1/day,1 次/天,连续注射10 d)引起大鼠双侧后肢50%机械刺激撤足阈值显著下降。如图2 A 所示,注射长春新碱后第4 d,同侧后肢的50% 机械刺激撤足阈值已由术前的25.67 g 降至6.52 g(Mann-WhitneyU检验,Z=-2.032,P=0.042),之后持续维持在比较低的水平,术后19 d 时仍然维持在低水平(Mann-WhitneyU检验,Z=-2.023,P=0.043),对侧后肢的50%机械刺激撤足阈值的变化与同侧后肢类似,说明腹腔注射长春新碱可引起大鼠双侧后肢长时间的机械痛觉过敏。
2.2 弥可保抑制长春新碱引起的脊髓背角病理性突触可塑性的发生
C 纤维介导的突触传递效率的增强与疼痛密切相关,其表现为脊髓背角C 纤维诱发场电位幅值的增大。如图2 B,我们发现与生理盐水组相比,长春新碱造模组大鼠在第19 d时(从第一次给长春新碱算起),脊髓背角C 纤维诱发场电位幅值在给予坐骨神经5 V 或10 V 单次电刺激时均显著增大。值得注意的是,坐骨神经给予5 V 的单次电刺激时,长春新碱造模组的大鼠可被诱发C 纤维诱发场电位(0.018 ± 0.008)mV,而生理盐水对照组、生理盐水+弥可保组和长春新碱+弥可保组几乎不能诱发C-纤维诱发场电位。如图2 C,当刺激强度从5 V 逐渐增大到12.5 V 时,同一强度的刺激在长春新碱组大鼠诱发的C 纤维诱发场电位的幅值明显大于生理盐水组。与生理盐水组相比,当刺激强度为12.5 V 时,长春新碱组大鼠诱发的C 纤维诱发场电位的幅值明显增大(Z=-2.371,P=0.016);而长春新碱+弥可保组大鼠C纤维诱发场电位幅值明显小于长春新碱组(Z=-2.611,P=0.008),单独给予弥可保组大鼠C 纤维诱发场电位幅值也明显小于长春新碱组(Z=-2.627,P=0.009)。以上结果说明长春新碱处理可能导致了C 纤维介导的突触传递效率的增强;而预防性给予弥可保能够缓解长春新碱的上述作用,单独使用弥可保也能降低C 纤维诱发场电位的幅值,说明弥可保可能对正常动物的基础痛阈也有影响。
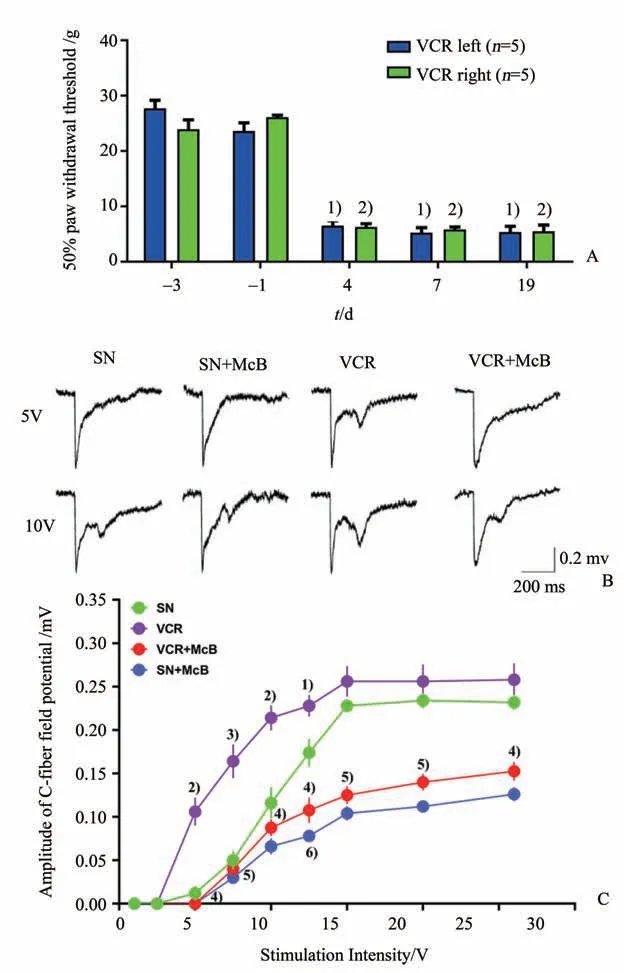
图2 腹腔注射长春新碱引起大鼠双侧后肢50%机械刺激撤足阈值显著下降和各组大鼠C纤维诱发场电位的记录情况Fig.2 Intraperitoneal injection of vincristine decreased the 50%paw withdrawal threshold in bilateral hindpaw in rats and the C-fiber evoked field potential recordings
2.3 弥可保抑制长春新碱引起的脊髓背角CGRP纤维芽生
如图3 所示,与生理盐水组(图3 A)相比,长春新碱给药第19 d 时,大鼠脊髓背角CGRP 阳性纤维的数目明显增多(图3 C,E;Z=-2.611,P=0.008),而给予弥可保(长春新碱给药前15 d 即开始给药,持续到最后一次长春新碱给药结束后9 d)可显著减少脊髓背角CGRP 的纤维数目(图3 D,E;Z=-2.611,P=0.008),提示抑制CGRP 纤维芽生可能是抑制脊髓背角C 纤维诱发电位长时程增强(long-term potentiation,LTP)的结构基础。弥可保慢性处理对正常大鼠脊髓背角CGRP 阳性纤维的数目无显著影响(图3 B,E;Z=-1.984,P=0.056)。
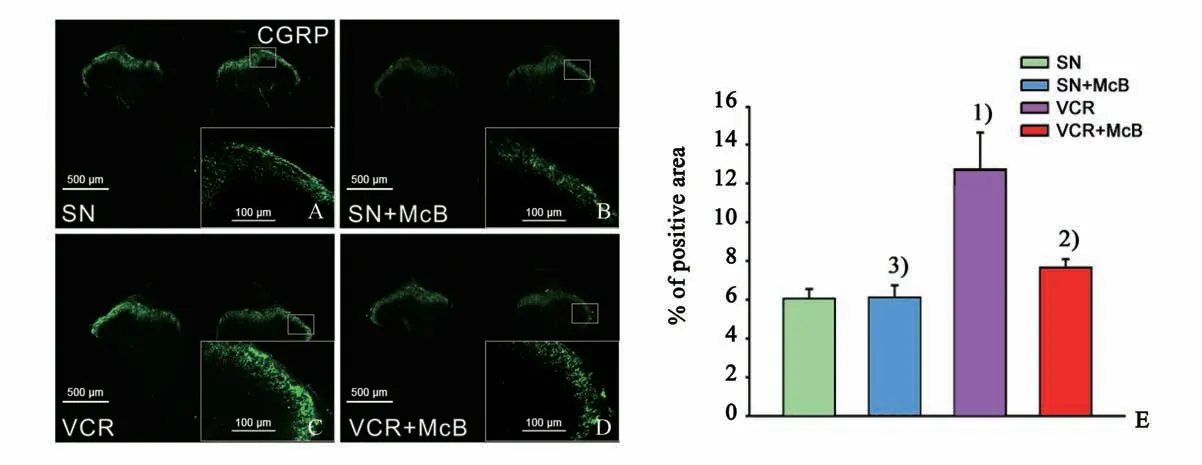
图3 腹腔注射弥可保可抑制长春新碱引起的脊髓背角CGRP阳性C纤维增多Fig.3 Intraperitoneal injection of McB inhibited the sprouting of CGRP positive C-fibers in the spinal dorsal horn induced by vincristine
2.4 弥可保防止长春新碱引起的脊髓背角NF-κB信号通路的激活
NF-κB 由p65 或p50 组成。p65 作为NF-κB 通路的主要部分,p-p65 表达的增加反映了NF-κB 的激活,NF-κB p65 激活之前需要磷酸化将其锚定到细胞核中特定的目标基因。NF-κB 也被报道可以调节一些致炎细胞因子的表达,从而在CIPN 中发挥重要作用[6]。因此我们用蛋白质印迹和免疫荧光方法观察了弥可保对脊髓背角p-p65 表达的影响,以此观察对NF-κB 通路激活的影响。Western Blot 结果表明,长春新碱造模组第14 d 时大鼠脊髓背 角p-p65 的表达 显著增加(图4A,F(4,20)=16.90,P<0.000 1;单因素方差分析继以Tukey′s 多重比较)。免疫荧光双染结果显示,p-p65 在NeuN标记的神经元(图4 B,D,K)、Iba1 标记的小胶质细胞(图4 E,G,K)及GFAP 标记的星型胶质细胞都有表达(图4 H,J,K)。与生理盐水组(图4 L)相比,长春新碱造模组大鼠在第19 d 时p-p65 表达增加(图4 M)。以上结果表明无论是神经元还是胶质细胞的NF-κB 通路在给予长春新碱后都被激活。进一步的实验表明,给予弥可保可显著减少长春新碱(第19 d)脊髓背角神经元及星形胶质细胞内p-p65 的表达(图4 K-N,Mann-WhitneyU检验,Z=-2.611,P=0.008),说明弥可保抑制了NFκB通路的激活。
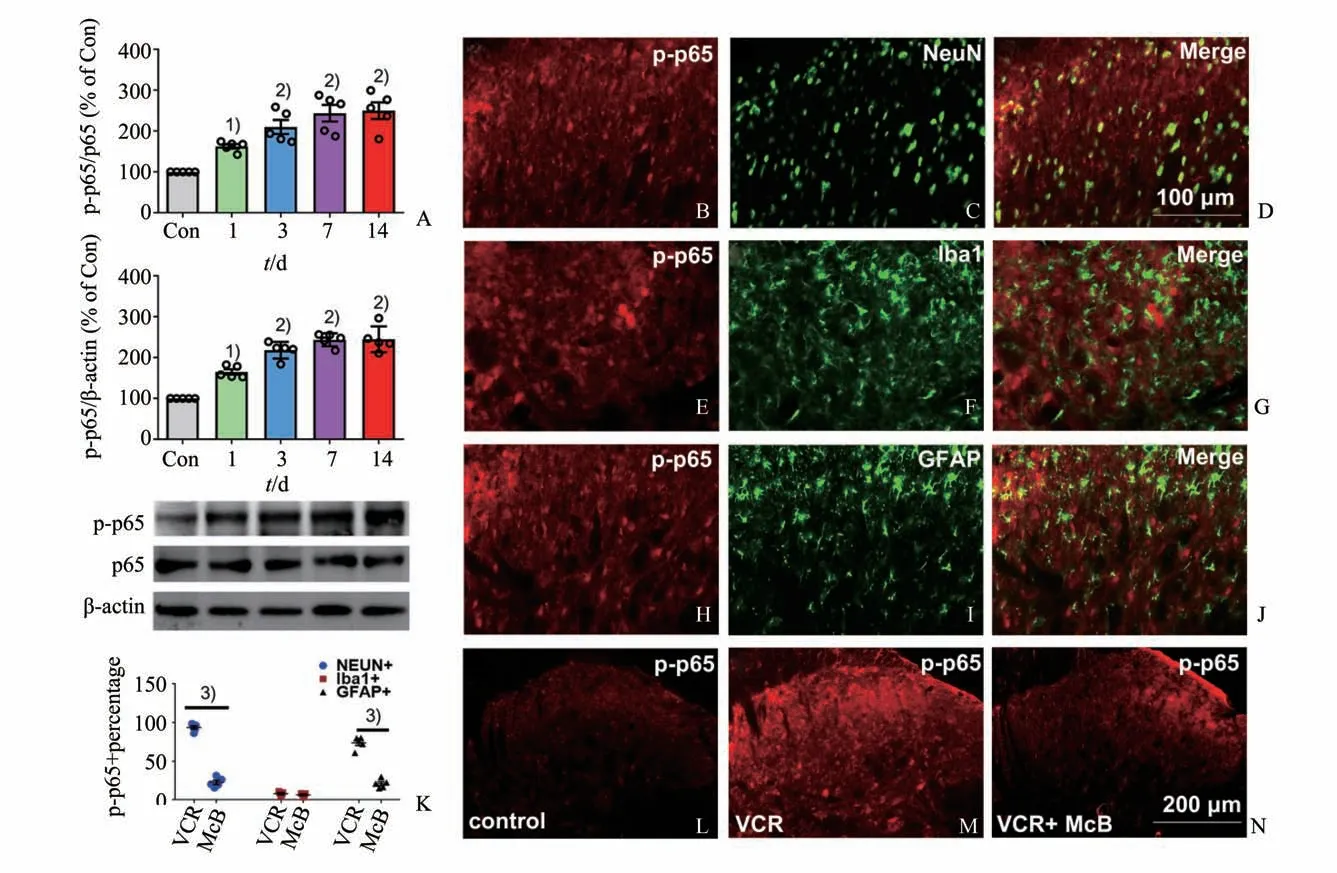
图4 腹腔注射弥可保可抑制长春新碱引起的脊髓背角p-p65表达增多Fig.4 Intraperitoneal injection of McB inhibited the up-regulation of p-p65 in the spinal dorsal horn induced by vincristine
IL-10 作为一种抗炎细胞因子,也可以抑制如IL-1β、TNF-α 和IL-6 等多种促炎细胞因子的产生[17]。我们观察到与生理盐水组相比(图5 A,D),长春新碱组给药后第19 d 时脊髓背角IL-10 的表达显著降低(图5 B,D,Mann-WhitneyU检验,Z=-2.611,P=0.009),而给予弥可保组大鼠其脊髓背角抗炎细胞因子IL-10的表达明显上调(图5 C,D,Mann-WhitneyU检验,Z=-2.402,P=0.0159)。在生理盐水组,免疫荧光双染显示IL-10 只表达在神经元(图5 E-G),在星形胶质细胞(图5 H-J)及小胶质细胞(图5 K-M)没有观察到IL-10的表达。
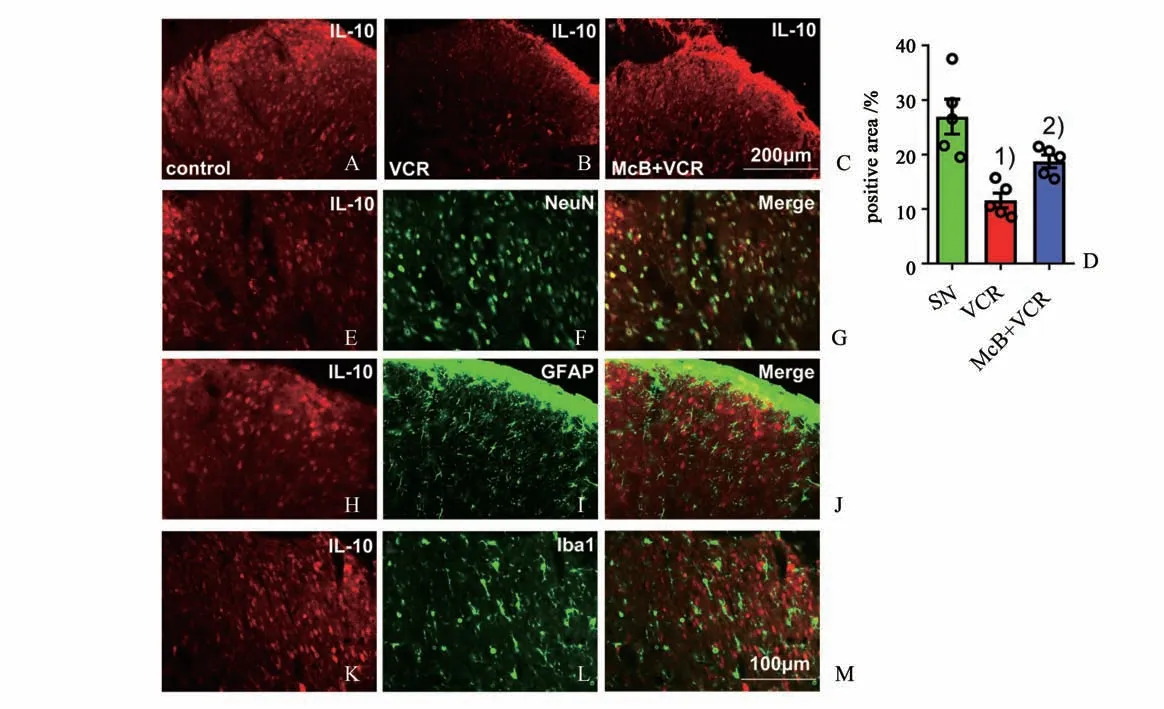
图5 预先给予弥可保可逆转长春新碱引起的脊髓背角IL-10表达减少Fig.5 Pretreatment with McB reversed the decrease of IL-10 in the spinal dorsal horn after vincristine treatment
3 讨论
在本研究中,我们发现预防性使用弥可保显著抑制了长春新碱诱导的脊髓背角C 纤维诱发场电位的增大。同时,弥可保抑制了脊髓背角CGRP 纤维的增多及NF-κB 通路的激活,并且增加了IL-10的表达。这些结果表明,弥可保通过抑制NF-κB通路的激活及纠正炎性细胞因子失衡防止中枢敏感化。
伤害性刺激可导致坐骨神经C 纤维与脊髓背角形成的突触传递效率显著增强。表现为脊髓背角神经元的兴奋性持续性增高,即形成了中枢敏感化,这可能是产生持续性疼痛的重要原因之一[18]。在本研究中,我们发现腹腔注射弥可保(维生素B12 的一种活性结构)可防止长春新碱诱导的脊髓背角C 纤维诱发电位LTP,提示弥可保可通过防止中枢敏感化起作用。除此之外,长春新碱处理也可引起外周敏感化[19],即导致初级传入纤维的自发活动(异位放电)[20],这也是引起神经病理性疼痛的关键因素[21]。最近的一项研究表明,弥可保可以显著地抑制CCD 大鼠背根神经节神经元的异位自发放电[22]。在本研究中,弥可保为腹腔注射,其可以通过血液循环到达全身各处组织,包括背根神经节,因此弥可保也有可能通过防止外周敏感化起作用。
化疗药物长春新碱的使用常常会导致病理性疼痛。传导慢性疼痛的C 纤维分为表达IB4非肽能纤维和表达CGRP 肽能两大类。我们之前的研究表明,长春新碱可引起脊髓背角CGRP 肽能C 纤维芽生,但不能引起IB4非肽能C 纤维芽生。这一变化可能是引起化疗后疼痛产生的重要原因[6]。在本研究中,我们发现弥可保可以防止CGRP 肽能C纤维芽生。这可能是弥可保减弱痛觉过敏的重要原因。
NF-κB 通路的激活可以使致炎细胞因子如TNF-α、IL-1β 和IL-6 等细胞因子的转录增多,其被证明参与了长春新碱诱导的痛觉过敏和痛觉异常的发展[23]。因此,减少炎性因子的生成被认为是防止病理性疼痛的有效手段,弥可保和天竺葵的合用已被报道可以通过减少TNF-α 的生成减缓泰素诱导的周围神经病变[24]。而我们之前的Western blot 实验也表明弥可保可减少TNF-α 在脊髓背角的表达量[14]。在本研究中,我们进一步发现,弥可保可以在脊髓背角抑制NF-κB p65的磷酸化,提示弥可保可通过抑制NF-κB 减少致炎细胞因子的表达。IL-10 是一种重要的抗炎细胞因子,在给予长春新碱后其表达显著下调,而弥可保则可以增加IL-10 的表达。与之一致的是,鞘内注射IL-10 基因治疗药物可以抑制泰素诱导的机械触诱发痛敏[25]。以上结果提示弥可保可能通过平衡致炎与抗炎细胞因子的表达,改善神经元生存的局部微环境,进而抑制神经病理性疼痛。
除了弥可保单独使用外,其通常还与维生素E等其他药物联合使用[26]。在2 型糖尿病引起的周围神经病变中,弥可保、α-硫辛酸和普瑞巴林的联合使用比单独使用普瑞巴林提供了更强的镇痛作用[27]。因此弥可保与其他药物联合使用可能比单独应用更加有效。
在本研究中,我们发现与生理盐水处理相比较,单独慢性给予弥可保可显著下调刺激C 纤维诱发场电位的幅度,说明弥可保本身对正常状态下C纤维与脊髓背角感觉细胞的突触传递可能有抑制作用,提示其对治疗急性疼痛可能也有效。但在之前的相关研究中,我们发现单独使用弥可保对正常大鼠对脊髓后角相关炎性指标IL-10 及TNF-α 的表达并无显著影响[13],在本研究中单独使用弥可保对正常大鼠CGRP 表达也无显著影响。综合判断,我们推测弥可保抑制正常情况下突触传递的功能可能不是通过抑制脊髓背角炎性指标所进行的。有研究报道高浓度弥可保可以稳定局部细胞膜内外离子通道,降低细胞膜内外离子浓度差,减少动作电位产生,从而抑制异位放电[22]。我们推测弥可保可能通过抑制突触前动作电位的发放进而抑制了C 纤维神经递质,包括谷氨酸和ATP 向脊髓背角的释放,具体的机制还需要进一步实验探讨。
综上所述,本研究证明预防性使用弥可保可显著抑制长春新碱诱导的脊髓背角C 纤维诱发场电位的增大。同时,弥可保抑制了脊髓背角CGRP 纤维的芽生及NF-κB 通路的激活,并且增加了IL-10的表达。
