甘肃陇东大豆孢囊线虫田间侵染动态
2022-04-04罗宁李惠霞刘永刚李焕宇杨顺义李金章石明明李金鸿
罗宁 李惠霞 刘永刚 李焕宇 杨顺义 李金章 石明明 李金鸿


摘要 为探讨甘肃陇东地区大豆孢囊线虫田间侵染动态,于2017年-2019年对大豆根系和根际土壤定期定点取样,采用漂浮法、贝曼漏斗法及酸性品红染色法对土样中和根组织内线虫进行分离计数,研究其繁殖数量与气温、降雨量的关系。结果表明,2017年土壤中2龄幼虫和新鲜孢囊的数量有3个明显高峰,大豆根组织内2~4龄幼虫均出现3个明显高峰,即发生3代,平均世代历期35 d。2018年-2019年土壤中2龄幼虫和新鲜孢囊均有4个明显高峰,大豆根组织内2~4龄幼虫亦出现4个明显高峰,即发生4代。第1代侵染数量最多,是主要危害世代。最后1代侵染数量极少且发育缓慢。在2017年-2019年生长季中,线虫繁殖数量与生长季平均温度具有相关性,与累积降雨量无相关性。
关键词 大豆; 大豆孢囊线虫; 侵染特征; 世代循环
中图分类号: S435.651
文獻标识码: A
DOI: 10.16688/j.zwbh.2021019
Abstract In order to explore the infection dynamics of soybean cyst nematode in the soybean field in east of Gansu province, soil and root samples were periodically collected from one fixed soybean block from 2017 to 2019. The nematodes were separated and counted under microscope by floating, Baermann funnel and acid fuchsin staining method, and the relationship between the reproduction and temperature and humidity were investigated. The results showed that there were three distinct number peaks of 2nd stage juvenile and new cysts in the soil, and three peaks of the emergence of 2nd, 3rd and 4th juveniles in soybean roots, which occurred in three generations and the average period of one generation was 35 d in 2017. From 2018 to 2019, there were four distinct number peaks of 2nd stage juvenile and new cysts in the soil and four peaks of the emergence of 2nd, 3rd and 4th juveniles in soybean roots, indicating that four generations occurred. The first generation had the largest number of infections, which was the main damage period and nematodes developed slowly in last generation. During the growing season from 2017 to 2019, the number of nematodes reproduction was correlated with the average temperature, but not with the accumulated rainfall.
Key words soybean; Heterodera glycines; infection characteristics; generation cycle
大豆孢囊线虫(soybean cyst nematode,SCN)属侧尾腺口纲垫刃目异皮科孢囊线虫属,是大豆生产中危害最严重的植物病原线虫[1]。目前,在美国、加拿大、波多黎各、海地、阿根廷、巴西、巴拉圭、哥伦比亚、俄罗斯、比利时、英国、意大利、日本、韩国、朝鲜、伊朗、印度尼西亚和埃及等国家大豆种植区普遍发生[25]。我国23个省(自治区,直辖市)已有大豆孢囊线虫分布[611],其中在黄淮海和东北大豆产区危害尤为严重,每年危害面积超过267万 hm2[12],随着大豆连作年限增加,病情逐年加重,严重者减产80%以上,甚至绝收。
据报道,大豆孢囊线虫理论上1年可以繁殖6代,具体世代数主要受土壤温度、寄主品种、播种时间和播种期等因素影响[13]。美国明尼苏达州大豆孢囊线虫1年发生3~4代[14]。俄罗斯阿穆尔州地区1年发生3代,平均历期27.3 d[15]。在我国,黑龙江省1年发生3代,第1代发生量最大[16],在山西、江苏和河南等省1年发生4代[6]。同一地区不同生长季,大豆孢囊线虫发生代数存在差异。研究显示,大豆孢囊线虫2013年在山东大豆上发生3代,2014年发生4代,第2代有明显世代重叠现象[1314]。北京地区,大豆孢囊线虫在地黄上1年发生5~6代[17]。目前,甘肃省大豆孢囊线虫的侵染动态尚无研究。
甘肃省是西北地区大豆主产区之一,年种植面积维持在13.33万~15.33万hm2,其中陇东地区种植面积最大,达10万hm2[18],本课题组调查发现,陇东地区大豆孢囊线虫发生严重,孢囊检出率为87.2%,已成为该地区大豆生产的主要限制因素[19]。
在大豆生长季,孢囊线虫的侵染次数与数量受多种因素影响,如寄主品种、气温和降雨量等均不同程度地影响大豆孢囊线虫群体繁殖[2022],另外耕作方法和土壤质地也会导致大豆孢囊线虫群体变化[2325]。我国幅员辽阔,大豆种植区跨度大,甘肃与黑龙江、江苏及山东等大豆产区气候条件、大豆播种期均不同,大豆孢囊线虫侵染规律与其他地区是否存在差异尚不明确。因此,本研究在2017年-2019年春播大豆生育期内对大豆孢囊线虫群体变化动态进行系统研究,探究气温、降雨对其繁殖的影响,以期明确该地区大豆孢囊线虫田间侵染规律,为大豆孢囊线虫有效防治提供依据。
1 材料与方法
1.1 供试品种
大豆‘中黄30号’(市售),中感大豆孢囊线虫病。
1.2 试验地概况
试验地位于甘肃省庆阳市宁县良平镇丰乐村(35°26′E,108°04′N),年平均氣温8.7℃,无霜期168 d,年均降水量565.9 mm,降雨多集中在7月-9月,季节分配不均,属于典型黄土高原半湿润偏干旱气候类型区[26]。试验地连续多年种植大豆,孢囊线虫基数较高,平均37.4个/100 g。分别于2017年5月1日、2018年5月2日和2019年5月5日播种大豆,播种后田间无人工灌溉,进行正常锄草和喷药管理。
1.3 取样时间和方法
2017年5月15日至9月23日内共取样14次,每间隔10 d左右取样1次。2018年5月13日至9月21日内共取样19次,2019年5月15日至9月20日内共取样19次,每间隔6~8 d取样1次。采用“ZigZag”取样法,每次随机选取10个样点,每个样点挖采大豆根系2~3株,保持根系完整,装入取样袋。每个样点挖取根围深度0~20 cm土壤作为1个小样,将10个小样装入塑料袋混合作为1个样品,约1 kg,带回实验室检测。
1.4 线虫的室内检测
1.4.1 土壤中线虫的分离
将土样混合均匀后称取100 g,采用漂浮法和贝曼漏斗法对孢囊和2龄幼虫(J2)进行定量分离[27],在60目筛上收集白雌虫和孢囊,解剖镜和显微镜下对孢囊和J2进行镜检和计数。
1.4.2 根组织中线虫的染色和计数
将混合样中的大豆根系清洗干净剪成2~3 cm小段并称重,采用酸性品红对根组织进行染色[4]。在显微镜下对染色的2龄、3龄(J3)、4龄(J4)幼虫和根组织内年轻雌虫进行镜检、计数和拍照。
1.4.3 孢囊线虫世代的划分标准
依据土壤中J2、新鲜孢囊出现的高峰数及其时间间隔长短,同时参考根组织中J2、J3、J4和年轻雌虫出现情况来确定大豆孢囊线虫的世代。
1.5 数据统计与分析
统计每100 g土壤中孢囊(白雌虫和新鲜孢囊)和J2的数量,以及1 g根组织内J2、J3、J4和年轻雌虫的数量,统计各虫态发育动态。从当地气象局获取当年气象数据,统计每个取样周期间平均气温、平均最低气温、平均最高气温和累积降雨量。采用Excel 2010和SPSS软件对各龄期群体动态、气温和降雨信息进行统计分析。
2 结果与分析
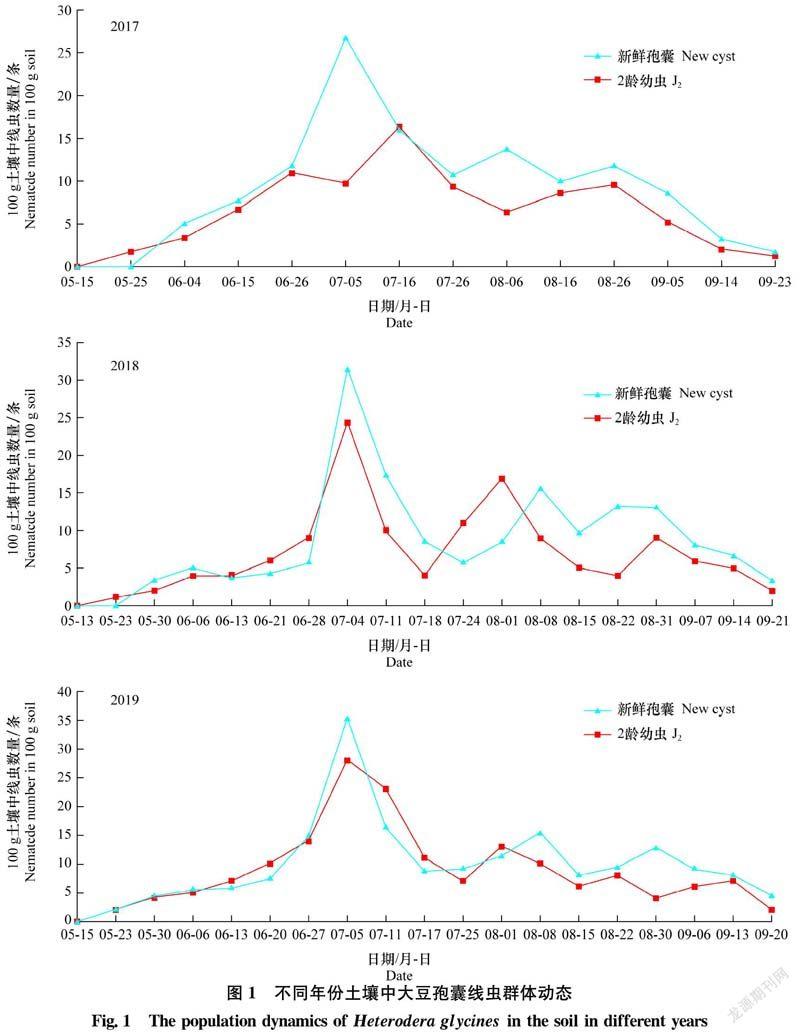
2.1 土壤中大豆孢囊线虫群体动态
2017年-2019年3个大豆生长季对土壤中孢囊线虫J2和新鲜孢囊群体密度变化情况的观察表明,100 g土壤中新鲜孢囊和J2数量的变化趋势较为吻合,有轻微起伏(图1)。2017年新鲜孢囊出现3个高峰,2018年-2019年新鲜孢囊出现3个明显高峰和1个不明显高峰。2017年-2019年新鲜孢囊数最高值分别出现在7月5日、7月4日和7月5日,100 g土中分别有26.7、31.0个和35.3个。9月中旬,新鲜孢囊数量持续减少,孢囊数量为1~5个。此外,在2018年6月6日发生高峰不明显,孢囊数仅为5个。
2017年-2019年,土壤中J2群体数量变化见图1。2017年土壤中J2数量共出现3个明显峰值,时间为6月26日、7月16日和8月26日。7月16日J2密度最大,100 g土中有16.3条。生长季后期,J2群体密度逐渐降低,数量极少,100 g土只有2条。2018年-2019年,土壤中J2数量均出现4个明显峰值,J2全年密度最大出现在7月4日和7月5日,为24.3条和28.0条,9月中旬土壤中J2发生水平低,仅为1~2条。
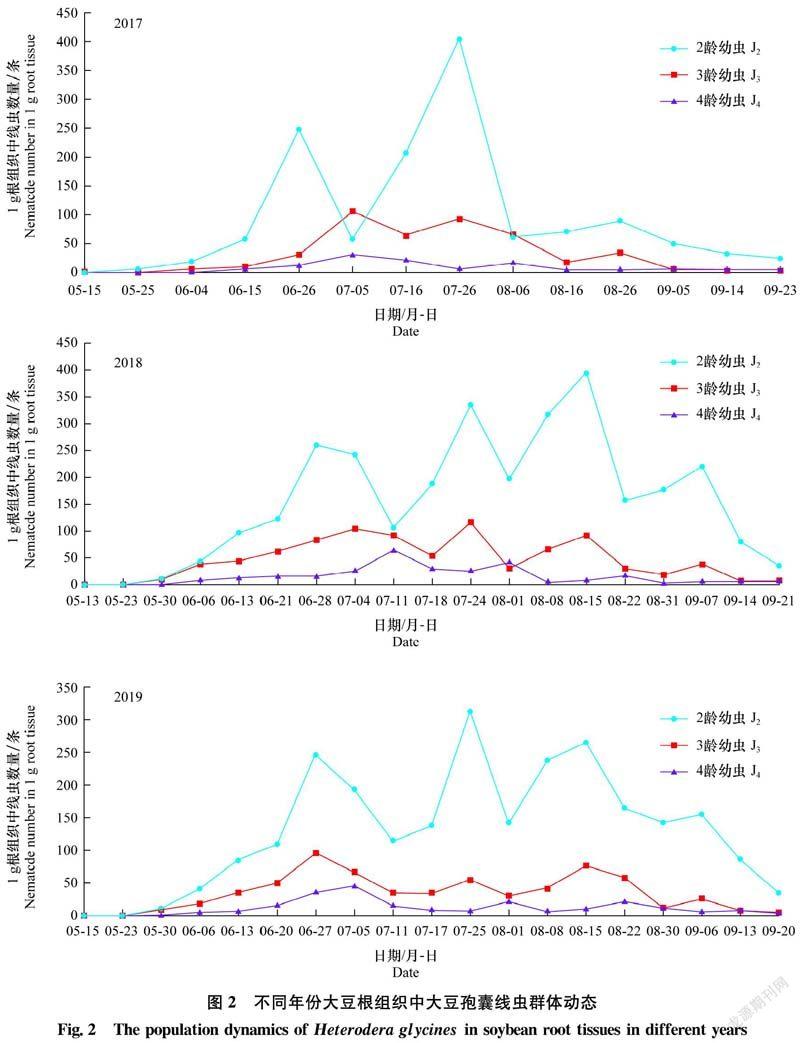
2.2 根组织中大豆孢囊线虫群体动态
2017年-2019年大豆根组织中大豆孢囊线虫 J2、J3和J4群体密度变化情况见图2。2017年J2密度有3个明显高峰,其中7月26日密度最大,1 g根系中有404条。J3和J4亦有3个明显高峰,密度最高为1 g根中有105条、31条。9月5日-14日根组织中J3和J4密度极小,1 g根中仅有3条。2018年和2019年,J2侵染数量均有4个明显高峰,分别在8月15日和7月25日J2数量最多,1 g根中分别有394条和312条。J3数量有4个明显高峰,在7月24日和6月27日数量最多,1 g根中分别有115条和95条,持续时间较短。J4数量有4个明显高峰,分别在7月11日和7月5日数量最多,分别为64条和45条。9月上旬以后根组织内虽然再次出现J3和J4,但密度极小,仅有2~4条。
2.3 大豆孢囊线虫的世代循环
2017年-2019年,从6月上旬至7月上旬,土壤中J2与新鲜孢囊发生水平低,结合根组织中J2侵入量、J3和J4(图3,b~e)形成量大幅增加可以推测基本完成第1代。第1代J2侵入速度慢,根组织中各龄幼虫发育迟缓,数量多,世代周期长,约42 d。
2017年7月5日至9月6日,根组织中J2、J3和J4均出现2个高峰,表明基本完成第2、3代,历期分别为32 d和30 d。第1代和第2代内各虫态发生高峰顺序与土壤中J2→根组织中J2、J3、J4→新鲜孢囊不相符,表明前2代存在较为明显的世代重叠现象。第3代根内J3、J4发育缓慢且数量极少,土壤中J2与新鲜孢囊数量呈下降趋势,说明第3代发育不完全。因此,2017年大豆孢囊线虫发生3代。
2018年和2019年,从7月上旬至8月上旬,土壤中J2、根组织中各龄幼虫与新鲜孢囊在不同时期发生不同程度的高峰,表明基本完成第2代,历期28 d,根组织中J2、J3、J4发育较慢、世代周期较长。第1代和第2代内各虫态发生高峰的顺序并不规律,表明前2代存在较为明显的世代重叠现象。第3代于8月下旬土壤中J2,根组织内J2、J3、J4数量达到高峰,且土壤中产生大量新鲜孢囊,该世代历期21 d基本完成,J2侵入速度快,根组织中幼虫发育快且世代发生比较整齐。直至9月中旬根组织内J2侵染数量达到高峰,但根内J3、J4发育减慢,数量极少,土壤中J2出现峰值,完成第4代,历期23 d。之后虽然在土壤中能检测出少数J2,但对根组织的侵染基本停止。因此,2018年和2019年大豆孢囊线虫发生4代。
每代根组织内J4膨大突出根部表皮,进一步发育为成熟雌虫(图3,f~g),雌虫在体内产卵。 后期雌虫死亡,体壁渐渐增厚,形成褐色、柠檬形的孢囊(图3h)脱落到土壤中。孢囊中的卵粒(图3a)存活时间长。由此可见,大豆孢囊线虫每年第1代侵染时间基本一致,但在6月下旬后,因田间温度和降雨量影响,大豆孢囊线虫侵入和发育速率发生变化,2018年和2019年第2、3、4代历期较短。
2.4 温度和降雨对大豆孢囊线虫繁殖的影响分析
该地区在大豆整个生长季无人工灌溉,因此气温是影响大豆孢囊线虫繁殖的主要因素。用SPSS软件做Spearman相关分析,结果表明,平均温度与J2、J3、J4数量的Spearman相关系数分别为0.747、0.737、0.608 (P<0.001),说明平均温度(℃)和线虫数量(条)存在正相关关系,即平均温度升高与线虫繁殖数量有关(表1)。而累积降雨量与J2、J3、J4数量的Spearman相关均为P>0.05,说明累积降雨量(mm)和线虫数量(条)不存在相关关系,可能和降雨量不均匀有关。
3 讨论
大豆孢囊线虫在20~28℃内随温度升高而发育速度加快,28℃为最适发育温度,高于33℃几乎不能发育[27]。在甘肃陇东,气象数据表明,2017年7月份最高气温超过30℃的高温日数多达22 d,平均最高气温29.0℃(21.8~38.5℃),持续时间较长,降雨量少,土壤呈半干旱状况,土壤中J2数量显著减少。2018年和2019年7月份最高气温超过30℃的高温日有8 d,平均最高气温27.5℃(20.1~32.0℃),降雨量较2017年适中,土壤中J2数量明显增加,这表明持续过高的温度和过低的土壤含水量抑制大豆孢囊线虫的侵染与生长发育。通过Spearman相关分析表明,线虫的繁殖数量与平均温度正相关,与降雨量无相关性,这与上述结果不一致,可能和最高气温持续期不同、降雨量分布不均匀有关,土壤中含水量过低或过高都会影响线虫的生长,导致线虫代数不完全和世代重叠现象出现,从而表现无相关性。降雨量和土壤含水量对大豆孢囊线虫影响复杂,除了干旱和潮湿的影响,还应分析水分胁迫强度及时间等[28],此部分内容有待更深层次研究。此外,每年9月中旬基本完成最后1代,根组织内各龄期幼虫数量少,土壤中成熟孢囊数量少、个体小,这可能是大豆生长季后期植株衰老、根组织木栓化和供给营养少所致。
刘维志[6]指出,在中国,大豆孢囊线虫1年可发生3~4代或6代。赵洪海等[13]和齐娜伟等[14]的研究表明,山东省大豆孢囊线虫在2013年发生3代,2014年发生4代,各世代平均历期30 d。美国明尼苏达州南部大豆孢囊线虫1年发生3~4代,从7月到11月,土壤中孢囊数量呈上升趋势。本研究表明,2017年甘肃陇东地区大豆孢囊线虫发生3代,各代平均历期较长,为35 d。2018年和2019年各代发育历期较短,平均为28 d,发生4代。本文报道大豆孢囊线虫世代平均历期和世代数与上述报道略有差异,说明大豆孢囊线虫世代历期与当地温度、品种抗性、土壤状况和田间管理措施密切相关[12, 29]。本研究发现,土壤中J2侵入点是新生伸长组织,在侧根和新生侧根中各龄期线虫数量多,主根中线虫数量随主根的生长而减少。在线虫大量孵化时期,一条新生须根中线虫侵染量最多达53条。
大豆孢囊线虫属于土传线虫病害,危害程度重,传播途径多,发病区域广。孢囊抗逆性较强,能够在干旱和低温条件下长期存活,防治极其困难[20, 30]。本研究结果表明,大豆孢囊线虫第1代发生量最大,是主要危害期,此时大豆根系脆弱、抵抗力差,容易被线虫侵染,因此应以防治第1代发生为主,控制第2、3代侵染为辅。本课题组田间试验表明,播种前,沟施0.5%阿维菌素颗粒剂和采用生物种衣剂菌线克SN101、SN102进行种子包衣处理均有一定防效[31]。大豆收獲后改种非寄主植物小麦、玉米、油菜和苜蓿等3~4年,每年可使土壤中孢囊基数减少20%左右[32]。
参考文献
[1] JONES J T, HAEGEMAN A, DANCHINE C J, et al. Top 10 plant parasitic nematodes in molecular plant pathology [J]. Molecular Plant Pathology, 2013, 14(9): 946961.
[2] 孙玉秋, 许艳丽, 李春杰, 等. 黑龙江省大豆胞囊线虫种群分布和卵密度研究[J]. 大豆科学, 2011, 30(2): 250253.
[3] MAAFI Z T, SALATI M, RIGGS R D. Distribution, population density, race and type determination of soybean cyst nematode, Heterodera glycines, in Iran [J]. Nematology, 2008, 10(6): 919924.
[4] 段玉玺. 植物线虫学[M]. 北京: 科学出版社, 2011.
[5] 陈井生. 黑龙江省大豆胞囊线虫毒力类型分析及品种抗性研究[D]. 沈阳: 沈阳农业大学, 2019.
[6] 刘维志. 植物病原线虫学[M]. 北京: 中国农业出版社, 2000.
[7] PENG Deliang, PENG Huan, WU Duqing, et al. First report of soybean cyst nematode (Heterodera glycines) on soybean from Gansu and Ningxia China [J]. Plant Disease, 2016, 100(1): 229.
[8] WANG Dong, DUAN Yuxi, WANG Yuanyuan, et al. First report of soybean cyst nematode, Heterodera glycines, on soybean from Guangxi, Guizhou and Jiangxi provinces, China [J]. Plant Disease, 2015, 99(6): 893.
[9] SHI Hongli, ZHENG Jingwu. First report of soybean cyst nematode (Heterodera glycines) on tobacco in Henan, Central China [J]. Plant Disease, 2013, 97(6): 852.
[10]ZHENG Jingwu, ZHANG Yan, CHEN Senyu, et al. First report of the soybean cyst nematode, Heterodera glycines, on soybean in Zhejiang, Eastern China [J]. Plant Disease, 2009, 93(3): 319.
[11]李克梅, 日孜旺古丽, 董艳秋. 新疆大豆孢囊线虫病的初步研究[J]. 植物保护, 2014, 40(2): 132134.
[12]宋美静, 朱晓峰, 王东, 等. 我国大豆主产区大豆胞囊线虫群体分布及致病性分化研究[J]. 大豆科学, 2016, 35(4): 630636.
[13]赵洪海, 丁海燕, 王凤龙. 大豆和烟草上大豆孢囊线虫田间侵染特征比较分析[J]. 中国农业科学, 2014, 47(22): 44174425.
[14]齐娜伟, 赵洪海. 2014年气象条件对大豆孢囊线虫田间侵染进程的影响分析[J]. 山东农业科学, 2015, 47(5): 8185.
[15]项鹏, 张武, 李宝华, 等. 俄罗斯阿穆尔州大豆胞囊线虫研究现状[J]. 黑龙江农业科学, 2018(8): 126127.
[16]刘晓英. 黑龙江省东部大豆孢囊线虫发生规律与防治技术研究[D]. 哈尔滨: 东北农业大学, 2003.
[17]陈金堂, 李知. 为害地黄的大豆孢囊线虫的初步研究[J]. 植物病理学报, 1981, 11(1): 3744.
[18]杨茹萍, 张国宏, 王立明, 等. 甘肃省大豆主产区产量性状及品质分析[J]. 大豆科学, 2013, 32(1): 5055.
[19]罗宁, 李惠霞, 郭静, 等. 甘肃省陇东南大豆孢囊线虫的发生和分布[J]. 植物保护, 2019, 43(3): 165169.
[20]李秀花, 马娟, 陈书龙, 等. 不同植物对大豆孢囊线虫种群的影响[J]. 河北农业科学, 2010, 14(8): 109111.
[21]孙晶双, 郑雅楠, 段玉玺, 等. 温度、光照和土壤含水量对大豆胞囊线虫休眠卵孵化的影响[J]. 大豆科学, 2009, 28(4): 690692.
[22]董秋月, 郑玉华, 赵洪海. 土壤水分对烟草孢囊线虫侵染和烟碱含量的影响[J]. 中国烟草科学,2015, 36(5): 6468.
[23]朱艳, 陈立杰, 段玉玺. 不同耕作方式对大豆胞囊线虫群体数量的影响[J]. 大豆科学, 2007, 26(2): 208212.
[24]靳学慧, 辛惠普, 郑雯, 等. 长期轮作和连作对土壤中大豆胞囊线虫数量的影响[J]. 中国油料作物学报, 2006, 28(2): 189193.
[25]许艳丽, 李春杰, 刘金波, 等. 东北黑土区重迎茬对大豆病虫害发生的影响[J]. 大豆科学, 2008, 27(3): 471474.
[26]路亚奇, 焦美龄, 曹彦超. 近35年甘肃庆阳黄土高原地带气候特征分析[J]. 中国农学通报, 2015, 31(29): 181190.
[27]刘维志. 植物线虫研究技术[M]. 沈阳: 辽宁科学技术出版社,1995.
[28]赵洪海, 梁晨, 许冠堂, 等. 土壤湿度对大豆孢囊线虫侵染和繁殖的影响[J]. 青岛农业大学学报(自然科学版), 2017, 34(1): 912.
[29]DUAN Yuxi, ZHENG Yanan, CHEN Lijie, et al. Effects of abiotic environmental factors on soybean cyst nematode [J]. Agricultural Sciences in China, 2009, 8(3): 317325.
[30]吴海燕, 段玉玺, 李秀侠. 大豆孢囊线虫的休眠[J]. 动物学报, 2006, 52(3): 498503.
[31]罗宁, 刘永刚, 李惠霞, 等. 不同杀线剂对大豆孢囊线虫病的防治效果[J]. 陜西农业科学, 2020, 66 (1): 1014.
[32]袁明. 黑龙江省西部大豆胞囊线虫病发生动态及防治对策[J]. 黑龙江农业科学, 2011(5): 4748.
(责任编辑:田 喆)
