基于优化MaxEnt模型的疣果匙荠在中国的适生区预测与分析
2022-04-04郭云霞王亚锋付志玺马蓿
郭云霞 王亚锋 付志玺 马蓿
摘要 本文优化了疣果匙荠分布数据筛选方式和MaxEnt软件参数设置,使用ENMTools剔除了冗余分布数据,调用R程序包Kuenm从1 240个不同参数组合的MaxEnt模型中筛选确定了最优参数。基于分布数据和参数优化的MaxEnt预测结果表明,疣果匙荠在中国的高度适生区和中度适生区分别占陆地总面积的9.4%和60.1%左右,温度和降水量是影响疣果匙荠分布的主要非生物因素。疣果匙荠在中国的入侵风险主要集中在4个方面,一是随植物引种传入扩散,二是国内既有发生区的扩大,三是随进口粮谷传入扩散,四是沿边境地区自然传入。
关键词 疣果匙荠; MaxEnt; 适生区; 参数优化
中图分类号: Q948
文献标识码: A
DOI: 10.16688/j.zwbh.2021148
Abstract The selection method of distribution data of Bunias orientalis and the parameter setting of MaxEnt software were optimized in this study. The redundant distribution data were eliminated by ENMTools, and the optimal parameters were selected from MaxEnt model with 1 240 different parameter combinations by using the R program package Kuenm. The prediction results with MaxEnt showed that the highly and moderately suitable areas accounted for about 9.4% and 60.1% of the total land area in China, respectively. Temperature and precipitation were the main abiotic factors affecting the distribution of B.orientalis. The invasion risks of B.orientalis in China were mainly concentrated on four aspects: spread accompanying the introduction of plant, expansion of domestic existing areas, spread accompanying the introduction of imported grain, and natural spread along the border areas.
Key words Bunias orientalis; MaxEnt; potential geographical distribution; optimization of parameter setting
疣果匙薺Bunias orientalis L. 是列入我国《进境植物检疫性有害生物名录》的入侵性杂草,属十字花科Brassicaceae,匙荠属Bunias,起源于高加索南部地区并在过去250多年里迅速扩张,现已遍布中欧和斯堪的那维亚半岛[1],欧洲东部和南部以及俄罗斯远东和西伯利亚地区、亚洲的中亚和西亚、北美洲的美国和加拿大也有分布,我国目前仅在东北、甘肃和北京有零星发现[25]。该物种生态适应性极强,能产生大量种子形成庞大的土壤种子库,可由小段残根或刈割残茬迅速再生,并可能借助化感作用抑制其他植物生长,产生有毒化学物质抵御植食性昆虫取食,从而迅速形成单一茂密群落,与本土植物争夺养分、阳光和传粉昆虫,对本地生态系统和生物多样性构成严重威胁[68]。此外,疣果匙荠还充当了某些重要植物病毒的天然越冬寄主,促进了植物病毒在田间的保存和流行[9]。我国出入境检疫部门曾多次从进口粮谷油料等货物中截获疣果匙荠籽实,仅2015年-2019年从进口油菜籽中截获疣果匙荠的次数就占同期该类货物中检疫性杂草截获总次数的9.75%[10],因此该入侵性杂草随进口货物传入我国的风险极高。对疣果匙荠在中国的适生区进行预测和分析,是开展定量风险评估的基础,能为控制预案的制定提供科学依据,有助于及时优化调整防控策略[1112],防止该有害生物在中国的入侵和扩散。
最大熵模型MaxEnt是一种基于物种已知分布信息并结合环境数据来预测未知分布情况的数理统计方法,该模型操作界面友好、建模程序相对简单,尤其是对样本数量和质量的耐受度高,预测结果优异,是近年来物种适生区预测领域最流行的模型之一[1315]。我们参考MaxEnt应用研究领域的最新进展,对疣果匙荠全球分布数据进行了科学处理,对软件参数作了优化,预测了疣果匙荠在中国的适生区分布,分析了疣果匙荠在中国的入侵风险。
1 材料与方法
1.1 软件与数据准备
1.1.1 软件
MaxEnt 3.4.1软件从https:∥biodiversityinformatics.amnh.org/open_source/maxent/网址下载;R 3.6.3软件从https:∥cran.r-project.org/网址下载;Kuenm 1.1.6程序包从https:∥github.com/marlonecobos/kuenm网址下载;ENMTools 1.0.4软件从https:∥github.com/danlwarren/ENMTools网址下载;ArcMap 10.4.1、SPSS 20.0和Google Earth 7.1由宜宾职业技术学院提供。
1.1.2 地图资料
从标准地图服务网(http:∥bzdt.ch.mnr.gov.cn/index.html)下载1∶20 000 000中国地图(审图号GS(2019)1822号)作分析底图。
1.1.3 原始数据
物种分布数据:采集疣果匙荠全球分布数据2 147条供后续处理。其中,从全球生物多样性信息平台(GBIF,https:∥www.gbif.org/)采集基于馆藏标本(preserved specimen)和文献记载(literature)的分布数据2 143条(无中国分布数据);从中国植物图像库(http:∥ppbc.iplant.cn/)采集北京植物园和北京药用植物园分布数据2条;从中国植物科学数据中心(https:∥www.plantplus.cn/cn)馆藏标本数据库采集中国甘肃和辽宁分布数据各1条[1621]。
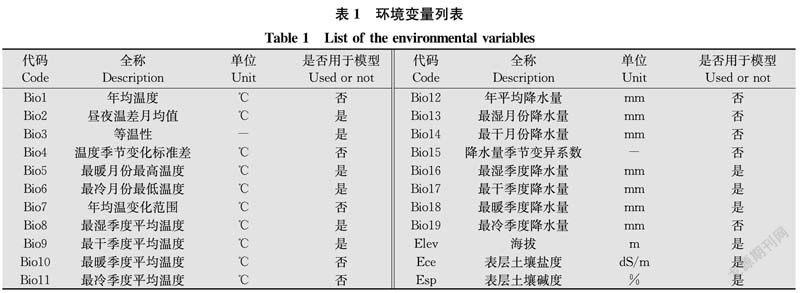
环境变量:选取22个环境变量供后续处理(表1)。1970年至2000年2.5 minutes精度的19个全球生物气候变量和1个全球海拔数据从世界气候网站(https:∥www.worldclim.org/)下载[1821];30 arc-second精度的全球土壤数据来源于联合国粮农组织世界土壤数据库V 1.2(http:∥www.fao.org/soils-portal/soil-survey/soil-maps-and-databases/harmonized-world-
soil-database-v12/en/),用ArcMap软件从中分别提取表层土壤碱度和盐度数据[2224]。
1.2 数据处理与参数优化
1.2.1 分布数据筛选方式优化
对部分仅标示采样地点而无经纬度坐标的物种分布信息,在Google Earth中匹配经纬度并转换为十进制数据。用ENMTools软件进行冗余分布数据剔除,取代常用的空间筛除法[13,25]。将分布数据导入ENMTools,以任一环境数据图层为背景,使用该软件的剔除重复点(removing duplicate occurrences)功能删去重复和位于同一环境变量数据栅格内的冗余数据,减少模型过拟合现象,提高预测质量。经筛选后保留参试分布数据1 014条,转换为CSV格式以便MaxEnt软件读取。
1.2.2 环境变量相关性分析与筛选
用SPSS对22个环境变量分组进行双变量相关性分析。若两个变量|r|≥0.8视为显著相关,则剔除生物学意义相对较小的1个变量,以减少模型过拟合[1920]。经筛选,最后保留9个全球生物气候变量、1个全球海拔数据、1个全球表层土壤碱度数据和1个全球表层土壤盐度数据共12个环境变量(表1)参加MaxEnt运算。用ArcMap统一12个环境变量的像元大小、范围和空间参考,并转换为ASC格式。
1.2.3 MaxEnt软件参数优化
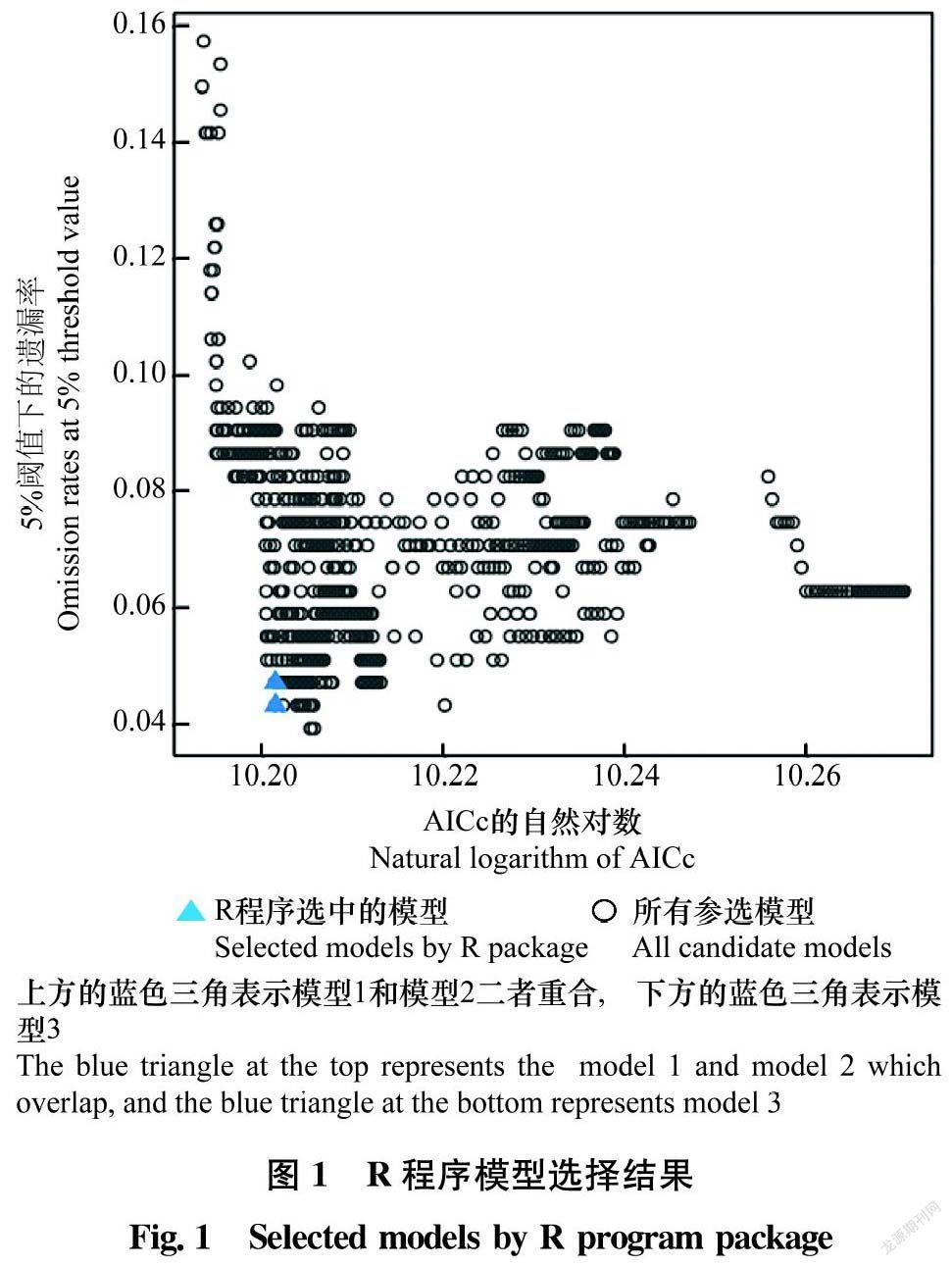
MaxEnt最重要的两个参数是特征组合(feature combination, FC)与正则化乘数(regularization multiplier, RM),對这两个参数的优化选择有助于显著提高模型的预测精度[1314,2629]。其中FC有5种可选项,分别是线性(linear,L)、 二次型(quadratic,Q)、乘积型(product,P)、阈值性(threshold,T)和片段化(hinge,H),可产生31种不同组合;RM参数一般设置为4以下,从0.1~4每间隔0.1设置1个RM值,共设置40个RM值[13]。调用R软件的Kuenm程序包,用MaxEnt进行1 240个不同参数模型(31种FC设置与40个RM值自由组合)的预测运算[27]。R软件先在所有参选模型中选出统计学上显著(significant models)且遗漏率(omission rates)≤5%的模型集,然后再根据赤池信息量准则(Akaike information criterion,AICc)选出Delta AICc值≤2的模型作为推荐模型[27,3031]。若R软件选出了1个以上推荐模型,则将其中Delta AICc值最小的1个模型作为最优模型。
1.2.4 疣果匙荠适生区预测
参照最优模型确定FC与RM的最佳配置,设定25%的参试样本作测试集,勾选切刀检验评估环境变量重要性,重复运算10次,将10次运算的平均值作为预测结果。MaxEnt软件将生成由一系列逻辑输出值(logistic output,LO)组成的全球地域图层ASC文件,LO数值范围在0~1之间,代表不同区域的适生指数(suitable index,SI)。根据疣果匙荠在全球的发生情况确定不同等级适生区划分的LO阈值。
2 结果与分析
2.1 最佳FC与RM参数
经R程序自动分析,所有1 240个参选模型统计学上均显著,并从中选出了3个推荐模型(图1),其中模型1的Delta AICc值最小(等于0),表明模型由已知分布区域向预测区域的移动性最好,且有效避免了模型过拟合[3031],为最优模型,相应的FC组合为Q+P+H,RM值为0.5。
2.2 疣果匙荠全球适生区预测结果
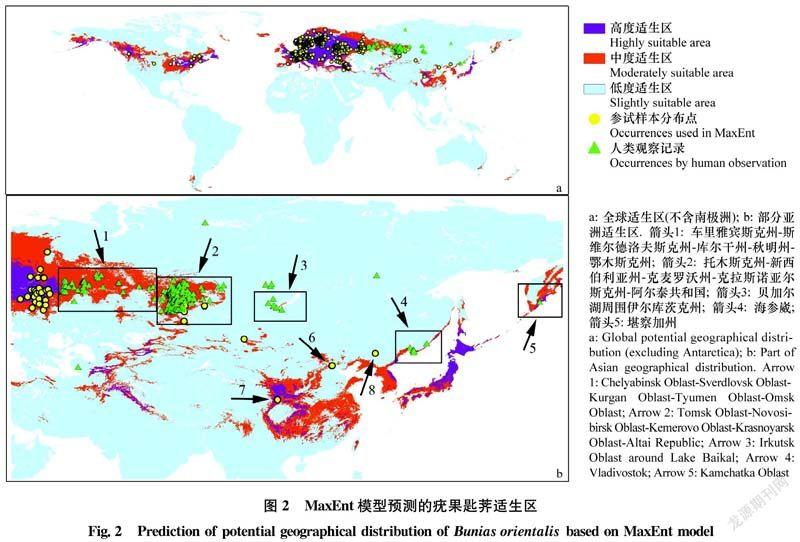
疣果匙荠全球适生区的LO值在0~0.810。参试样本分布点多集中在欧洲(图2),因此我们主要以欧洲区域为参考,将瑞典和芬兰南部、爱沙尼亚、法国、德国、俄罗斯西部等参试样本点密集连片的地域划分为高度适生区,LO值在0.155~0.810;将挪威西北部、瑞典西北部等参试样本点相对分散但仍有多个发生点的地域划分为中度适生区,LO值在0.022~0.155;其余只有零星发生或无发生记录的区域为低度适生区,LO值在0~0.022(图2a)。全球其他适生区域参照欧洲阈值划分。
为了直观评估预测效果,又从GBIF中选取了未用作参试样本、基于人类观察记录(human observation)的俄罗斯疣果匙荠分布数据250条(图1,绿色三角标识),用于验证MaxEnt预测结果在参试样本分布区域外的适用性。250个测试点的分布和评测情况如下(图2b): 1)车里雅宾斯克州斯维尔德洛夫斯克州库尔干州秋明州鄂木斯克州(箭头1),该区域无参试样本点,但预测结果显示沿着这5州有一条中度适生区分布带,经评测该区域确有超过50条人类观察的发生记录; 2)托木斯克州新西伯利亚州克麦罗沃州克拉斯诺亚尔斯克州阿尔泰共和国(箭头2),该区域仅有5个参试样本,但MaxEnt预测该区域存在部分高度适生区和大片中度适生区,经评测该区域有超过160条人类发现记录,且比较密集和连续; 3)贝加尔湖周围伊尔库茨克州(箭头3)、海参崴(箭头4)和堪察加州(箭头5),该3处均无参试样本分布,但MaxEnt预测出其存在中度或高度适生区,且被人类观察记录所支持。这在一定程度上印证了本文预测结果的准确性。
2.3 影响MaxEnt預测的主要环境变量分析
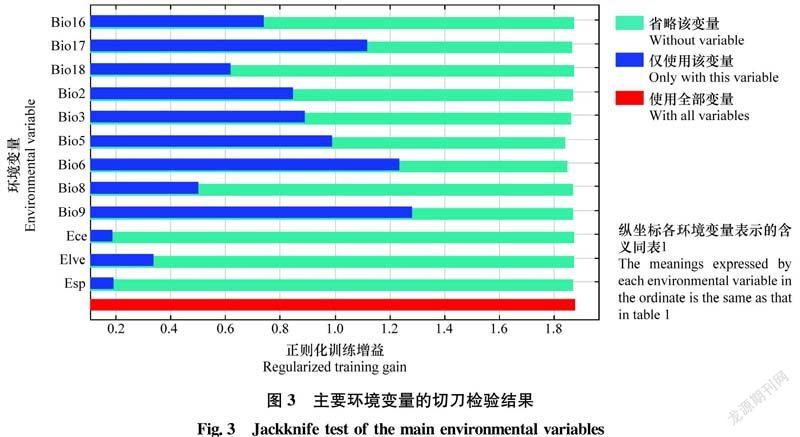
在切刀检验中,依次剔除每个变量时,模型增益并未显著减少,因此各变量包含的信息基本相同。但单独使用各个变量时,模型增益差异较显著,其中增益最多、也最重要的4个变量依次是Bio9(最干季度平均温度)、Bio6(最冷月份最低温度)、Bio17(最干季度降水量)和Bio5(最暖月份最高温度),表明温度和降水量是影响模型预测结果和疣果匙荠分布的主要非生物因素(图3)。
疣果匙荠高度适生区最干季度平均温度约在-14.1~24.3℃,最冷月份最低温度约在-21.7~5.4℃,最暖月份最高温度约在11.4~30.7℃,最干季度降水量约在2~550 mm;中度适生区最干季度平均温度约在-19.8~26.4℃,最冷月份最低温度约在-26.8~10.5℃,最暖月份最高温度约在5.9~34.1℃,最干季度降水量约在0~813 mm。根据植物生理学相关原理推测,低于-26.8℃或高于34.1℃有可能会对疣果匙荠造成低温或高温胁迫,从而影响其分布。疣果匙荠有一定的抗旱能力,最干季度降雨量很少时也可以维持正常生长,但对湿害或涝害的抗性不强,降雨量过多的地区对疣果匙荠适生性降低,可能因为水分过多引起含氮量减少和氧气缺乏,限制了植物的生长和代谢活动[22,32]。
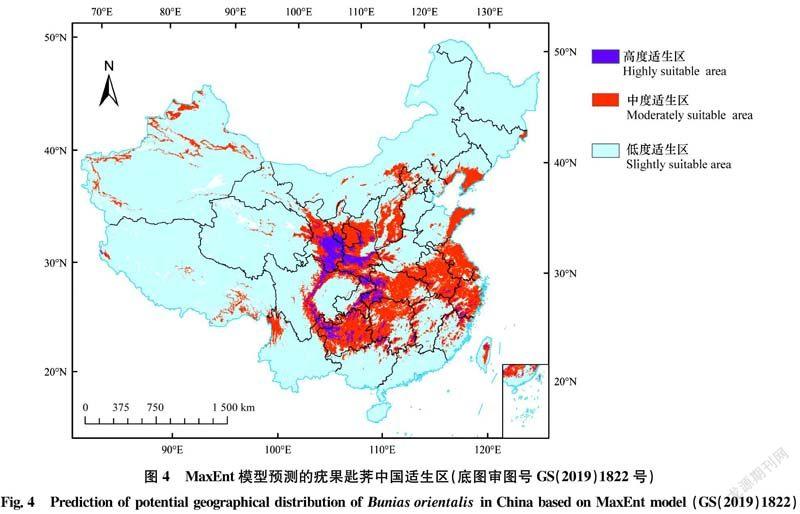
2.4 疣果匙荠在中国的适生区分析
疣果匙荠在中国的高度适生区占中国陆地总面积的9.4%左右,约90.24万 km2,主要分为4块(图4):一是毗邻的甘肃东南部、陕西中南部与宁夏南部部分区域;二是毗邻的陕西南部、重庆东北和东南部及湖北西部部分区域;三是沿四川东部高原山地和西部盆地的分界区域、云南东北部和贵州西部的部分区域;四是其余小块区域。
中度适生区约占中国陆地总面积的60.1%,约576.96万km2,是疣果匙荠在中国的主要适生区类型(图4),也可根据地理方位划为4大块:一是中部区块,主要包括毗连的甘肃东南半部、宁夏南半部、陕西大部、四川沿东部高原山地和西部盆地的分界线,以及云南东北和西北部、重庆东北与东南部、湖北大部、贵州大部及湖南西北半部;二是东部区块,主要包括河南东南部、安徽大部、江苏大部、浙江大部、上海、台湾中部,以及山东东部;三是东北部区块,主要包括吉林东部小块和辽宁南部、北京和河北部分地区、内蒙古中部和山西大部;四是其他相对分散的分布区域。
结合疣果匙荠在中国的发生记录,我们分析认为有4方面的疣果匙荠入侵风险应该被重点关注。
一是随植物引种传入扩散。境外引种是检疫性有害生物传入的主要渠道之一[3334]。中国植物图像库中疣果匙荠的拍摄地集中于北京植物园和北京药用植物园附近(图2b,箭头6)。类似地,同为检疫性杂草的齿裂大戟Euphorbia dentata Michaux最早于1976年在北京东北旺药用植物种植场发现,目前已定殖于北京且繁殖很快,推测是由园林植物引种带入[33,35]。虽然国家对植物引种制定了相应管理规定,但由于部分引种单位检疫风险意识不强、监管部门职责分散等原因,在实际施行中仍然存在一些实际困难[34]。
二是国内既有发生区的扩大。北京的疣果匙荠中度适生区以河北山西为纽带与中部的高、中度适生区连为一体,一旦疣果匙荠由北京扩散到达陕甘宁高度适生区并定殖泛滥之后,将有可能继续沿四川、湖北和重庆的高、中度适生区向贵州、湖南、安徽、江苏、浙江等连片的中度适生区入侵,造成难以控制的被动局面。以疣果匙荠在俄罗斯的发生为例,从车里雅宾斯克州到鄂木斯克州之间的中度适生区即充当了两个高度适生区连接的纽带(图2b,箭头1)。另外,数据库中有1条1959年采自甘肃武都(图2b,箭头7)的标本信息,但除此之外再无该区域的发生记录,因此存在两种可能,一是该区域少量发生时即被清除,二是目前该区域有发生但因地理位置较偏僻等原因而鲜见报道。若属后者,则存在极大扩散风险,因该发生点地处高度适生区中心地带,极易由此向全国高、中度适生区扩散。东北分布点(图2b,箭头8)虽多处于低度适生区范围,但也应防止其分布扩大并沿北京河北山西一线向中部和东部扩散。
三是随进口粮谷传入扩散。全国44%以上的进口粮食指定口岸(指定监管场地)分布在疣果匙荠高、中度适生区,尤其是辽宁、山东、江苏、浙江等地的指定口岸是进口粮食入境的主要集散点,很多内地企业加工的进口粮均是先在上述口岸卸船,然后经由水运(长江航运)或陆运(铁路和公路)运抵指定加工厂。进口粮食中极易携带疣果匙荠籽实,且易随粮食的接卸、运输、加工过程撒漏而扩散。加之进口粮食运输距离跨度大、加工地点分散偏僻,即使疣果匙荠在上述地区定殖也很难及时发现,给防控增加了一定难度。
四是沿边境地区自然传入。俄罗斯托木斯克州、新西伯利亚州、阿尔泰共和国等区域的密集发生点(图2b,箭头2)有可能通过新疆的中度适生区传入我国,海参崴的发生点(图2b,箭头4)也有可能传入我国东北。
3 討论
目前多数基于MaxEnt的物种适生区预测均采用了软件默认参数设置,未根据样本实际情况作相应调整和比较,然而样本处理和参数设置对模型预测结果影响很大[14,1621,2628]。
MaxEnt模型的一个基本假设是,所有目标区域都已被系统或随机取样, 然而在实践中,由于成本等原因限制,样本通常取自研究人员容易调查的区域,因此不可避免地导致某些区域样本量过于密集,引起取样偏差[13,31,36],导致MaxEnt模型过拟合,从而降低模型的预测能力[28,31,3638]。常见校正取样偏差的方法是空间筛除法,即在一定的地理半径距离内剔除过近的分布点,但该方法存在一个明显缺陷,就是忽略了分布数据与环境因子的相关性并且空间范围标准难以确定。在本文中,我们引入了一个环境生态位模型分析软件ENMTools[25]。该软件的突出优点是能将物种分布数据与环境数据关联起来进行匹配分析,在同一环境数据栅格内(即相近生态位)只保留一个有效分布数据,极大降低了分布数据过度集中导致的模型过拟合。
MaxEnt软件有多个可用的参数设置,确定最佳参数时应该考虑到统计显著性、预测能力和模型复杂性3个方面,因而需要通过模型校准找到最佳的参数组合,使预测结果最大趋近于物种的客观适生状况[2627,2930]。然而,进行人工校准非常耗时(通常需要几百上千次预测分析),甚至于难以实现[27]。在本研究中,我们参考近年最新研究进展,调用了一个名为“Kuenm”的R语言程序包实现了对MaxEnt模型重要参数的自动化校准和评估,从1 240种参数组合中选出了最优设置,进一步提高了适生区预测结果的可信度。我们创新性地使用参试样本分布点之外的分布数据评测了模型预测结果,更直观地展现了模型的预测精度。此外,国内外文献中对有害生物的适生区分级标准并不统一,从数级到十几级不等,国内运用比较多的是“高度适生区、中度适生区、低度适生区和非适生区”的4级分区法。然而,多数植物尤其是入侵性植物通常都具有高度的环境适应性与表型可塑性,存在较大的生存定殖潜力,因此我们认为,“非适生区”或“不适生区”的概念过于绝对化,不太符合植物生理学相关原理,容易导致植保工作者对该区域防控意识的麻痹和防控措施的松懈,且过细的分级对实际防控的指导意义不是很大,因而在本文中我们根据疣果匙荠在国外的分布传播情况,将其适生区分为3级,以期更直观地展示防控重点,指导防控预案制定。
在疣果匙荠的防控中,应积极贯彻“预防为主,综合防治”的植保方针。一是应加大对该杂草防控知识的宣教,使基层植保工作者和政府监管人员充分认识疣果匙荠入侵危害,掌握识别要点,并对目前已有发生点的区域做全面的调查和了解,及时铲除出现的植株并持续跟踪防除效果;二是应加强外来入侵杂草监测力度,尤其是对进口粮食指定口岸、指定加工厂,运输沿线周边高风险区域,以及植物引种园区、与有害生物密集发生点毗邻的地区等,开展有规律的常态化监测,确保早发现、早防除;三是监管部门应密切协作,加强对进口粮食口岸接卸、运输和加工储存、植物引种栽培等过程的监管,确保生产中的生物安全风险得到有效控制,防止疣果匙荠通过上述途径逃逸扩散,切实维护国内农林业生产和生态环境安全。
参考文献
[1] KOCH M A, MICHLING F, WALTHER A, et al. Early-mid Pleistocene genetic differentiation and range expansions as exemplified by invasive Eurasian Bunias orientalis (Brassicaceae) indicates the Caucasus as key region [J/OL]. Scientific Reports, 2017, 7(1): 16764. DOI: 10.1038/s41598-017-17085-8.
[2] 周太炎, 郭荣麟, 蓝永珍, 等. 中国植物志: 第33卷[M]. 北京: 科学出版社, 1987: 117.
[3] 傅沛云. 东北植物检索表[M]. 2版. 北京: 科学出版社, 1995: 242, 247.
[4] 中国科学院植物研究所. 中国高等植物图鉴: 第二册[M]. 北京: 科学出版社, 2016: 46.
[5] 印丽萍. 中国进境植物检疫性有害生物杂草卷[M]. 北京: 中国农业出版社, 2018: 34.
[6] KIETYK P. Distribution pattern of the invasive alien plant Bunias orientalis in Rów Podtatrzański trench, north of the Tatra Mts, Poland [J]. Biologia, 2014, 69(3): 323331.
[7] PATAMSYTE· J, NAUGEMYS D, C·EˇSNIENE· T, et al. Evaluation and comparison of the genetic structure of Bunias orientalis populations in their native range and two non-native ranges [J]. Plant Ecology, 2017, 219(1): 101114.
[8] DIETZ H, STEINLEIN T, WINTERHALTER P, et al. Role of allelopathy as a possible factor associated with the rising dominance of Bunias orientalis L. (Brassicaceae) in some native plant assemblages [J]. Journal of Chemical Ecology, 1996, 22(10): 17971811.
[9] KOBYKO T, MAJ Z, GAJEWSKI Z. Bunias orientalis L.as a natural overwintering host of Turnip mosaic virus [J]. Acta Agrobotanica, 2009, 62(2): 149154.
[10]袁俊杰, 盧乃会, 魏霜, 等. 2005-2009年进境油菜籽携带杂草疫情分析[J]. 中国植保导刊, 2020, 40(12): 8689.
[11]李志红, 秦誉嘉. 有害生物风险分析定量评估模型及其比较[J]. 植物保护, 2018, 44(5): 134145.
[12]万方浩, 侯有明, 蒋明星. 入侵生物学[M]. 北京: 科学出版社, 2015: 117126.
[13]孔维尧, 李欣海, 邹红菲. 最大熵模型在物种分布预测中的优化[J]. 应用生态学报, 2019, 30(6): 21162128.
[14]朱耿平, 乔慧捷. MaxEnt模型复杂度对物种潜在分布区预测的影响[J]. 生物多样性, 2016, 24(10): 11891196.
[15]PHILLIPS S J, ANDERSON R P, DUDK M, et al. Opening the black box: an open-source release of MaxEnt [J]. Ecography, 2017, 40(7): 887893.
[16]郭燕青, 史梦竹, 李建宇, 等. 基于MaxEnt模型的假臭草潜在分布区预测[J]. 热带亚热带植物学报, 2019, 27(3): 250260.
[17]林伟, 徐淼锋, 权永兵, 等. 基于MaxEnt模型的草地贪夜蛾适生性分析[J]. 植物检疫, 2019, 33(4): 6973.
[18]柳晓燕, 李俊生, 赵彩云, 等. 基于MaxEnt模型和ArcGIS预测豚草在中国的潜在适生区[J]. 植物保护学报, 2016, 43(6): 10411048.
[19]YAN Huyong, FENG Lei, ZHAO Yufei, et al. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt [J/OL]. Global Ecology and Conservation, 2020, 21: e00856. DOI: 10.1016/j.gecco.2019.e00856.
[20]XU Danping, ZHUO Zhihang, WANG Rulin, et al. Modeling the distribution of Zanthoxylum armatum in China with MaxEnt modeling [J/OL]. Global Ecology and Conservation, 2019, 19: e00691. DOI: 10.1016/j.gecco.2019.e00691.
[21]WEI Bo, WANG Rulin, HOU Kai, et al. Predicting the current and future cultivation regions of Carthamus tinctorius L.using MaxEnt model under climate change in China [J/OL]. Global Ecology and Conservation, 2018, 16: e00477. DOI: 10.1016/j.gecco.2018.e00477.
[22]GONG Zhizhong, XIONG Liming, SHI Huazhong, et al. Plant abiotic stress response and nutrient use efficiency [J]. Science China Life Sciences, 2020, 63(5): 635674.
[23]胡亮, 李鸣光, 韦萍萍. 入侵藤本薇甘菊的耐盐能力[J]. 生态环境学报, 2014, 23(1): 715.
[24]张风娟, 李继泉, 徐兴友, 等. 环境因子对黄顶菊种子萌发的影响[J]. 生态学报, 2009, 29(4): 19471953.
[25]WARREN D L, GLOR R E, MICHAEL T. ENMTools: a toolbox for comparative studies of environmental niche models [J]. Ecography, 2010, 33: 607611.
[26]RADOSAVLJEVIC A, ANDERSON R P. Making better MaxEnt models of species distributions: complexity, overfitting and evaluation [J]. Journal of Biogeography, 2014, 41: 629643.
[27]COBOS M E, PETERSON A T, BARVE N, et al. Kuenm: an R package for detailed development of ecological niche models using MaxEnt [J/OL]. Peer J, 2019, 7: e6281. DOI: 10.7717/peerj.6281.
[28]MEROW C, SMITH M J, SILANDER J A. A practical guide to MaxEnt for modeling species’ distributions: what it does, and why inputs and settings matter [J]. Ecography, 2013, 36: 10581069.
[29]ZENG Yiwen, LOW B W, YEO D C J. Novel methods to select environmental variables in MaxEnt: A case study using invasive crayfish [J]. Ecological Modelling, 2016, 341: 513.
[30]WARREN D L, SEIFERT S N. Ecological niche modeling in MaxEnt: the importance of model complexity and the performance of model selection criteria [J]. Ecological Applications, 2011, 21(2): 335342.
[31]WARREN D L, WRIGHT A N, SEIFERT S N, et al. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern [J]. Diversity and Distributions, 2014, 20: 334343.
[32]毛自朝. 植物生理学[M]. 武汉: 华中科技大学出版社, 2017: 360398.
[33]刘全儒, 于明, 周云龙. 北京地区外来入侵植物的初步研究[J]. 北京师范大学学报(自然科学版), 2002, 38(3): 399404.
[34]杨光, 吴新华, 顾忠盈. 对加强境外植物引种管理工作的思考和建议[J]. 植物检疫, 2005, 19(2): 121123.
[35]马金双, 程用谦. 中国植物志: 第44卷第3分册[M]. 北京: 科学出版社, 1997: 68.
[36]KRAMER-SCHADT S, NIEDBALLA J, PILGRIM J D, et al. The importance of correcting for sampling bias in MaxEnt species distribution models [J]. Diversity and Distributions, 2013, 19(11): 13661379.
[37]PHILLIPS S J, DUDK M. Modeling of species distributions with MaxEnt new extensions and a comprehensive evaluation [J]. Ecography, 2008, 31(2): 161175.
[38]ELITH J, PHILLIPS S J, HASTIE T, et al. A statistical explanation of MaxEnt for ecologists [J]. Diversity and Distributions, 2011, 17(1): 4357.
(責任编辑:杨明丽)
