农家品种‘白大头’及其衍生系‘天00127’苗期抗条锈性遗传分析
2022-04-04张静贾秋珍曹世勤孙振宇王万军黄瑾张勃王晓明
张静 贾秋珍 曹世勤 孙振宇 王万军 黄瑾 张勃 王晓明

摘要 2019年在甘肃省农业科学院植物保护研究所兰州温室,对抗条锈性优异的农家品种‘白大头’及其衍生系‘天00127’与感病品种‘铭贤169’杂交组合的P1、P2、F1、F2和BC1代材料,苗期分别人工接种条锈菌主要流行小种CYR34、CYR32的单孢菌系,进行抗病性遗传分析,结果表明:供试亲本‘白大头’和‘天00127’对CYR34和CYR32均表现免疫。各世代材料中,‘铭贤169’/‘白大头’组合与‘铭贤169’/‘天00127’组合的F1代对供试小种均表现免疫或近免疫;F2代植株表现抗感分离,其中‘铭贤169’/‘白大头’组合对CYR34符合1R∶3S的理论比值,对CYR32符合3R∶1S的理论比值;BC1代植株对CYR34表现全感,对CYR32符合1R∶1S的理论比值;‘铭贤169’/‘天00127’组合对CYR34和CYR32均符合3R∶1S的理论比值;BC1代植株抗感分离符合1R∶1S的理论比值。表明供试材料‘白大头’对CYR34和CYR32的抗性分别由1对隐性和1对显性抗性基因控制,‘天00127’对CYR34、CYR32的苗期抗性均由1对显性抗性基因控制。推测‘天00127’对CYR32的抗病基因可能来源于亲本农家品种‘白大头’或‘85-173-4’。
关键词 遗传分析; 苗期; 条锈病; 白大头; 天00127
中图分类号: S435.121.42
文献标识码: A
DOI: 10.16688/j.zwbh.2021032
Abstract The seedling resistance of wheat landrace ‘Baidatou’ and its derivative ‘Tian 00127’ by crossing with the susceptible variety ‘Mingxian 169’ and their offspring plants including F1, F2, and BC1 to CYR34, CYR32 of Puccinia striiformis f.sp. tritici (Pst) were evaluated in the Lanzhou greenhouse, Institute of Plant Protection, Gansu Academy of Agricultural Sciences (IPP, GAAS) in 2019. The results showed that the landrace ‘Baidatou’ and its derivative ‘Tian 00127’ were immune to CYR34 and CRY32. The F1 plants derived from the cross of ‘Mingxian 169’ with ‘Baidatou’ and the cross of ‘Mingxian 169’ with ‘Tian 00127’ were both immune or closely immune to CYR34 and CYR32. The segregation ratio for the cross between ‘Mingxian 169’ and ‘Baidatou’ accorded with the expected ratio of 1R∶3S to CYR34, and 3R∶1S to CYR32 in F2 plants; and all BC1 plants were susceptible to CYR34, but with a ratio of 1R∶1S to CYR32. The segregation ratio for the cross between ‘Mingxian 169’ and ‘Tian 00127’ accorded with the expected ratio of 3R∶1S to CYR34 and CYR32 in F2 plants, and BC1 plants showed an expected segregation ratio 1R∶1S. The results indicated that the resistance of wheat landrace ‘Baidatou’ was controlled by one recessive gene to CYR34, by one dominant gene to CYR32. The resistance of wheat ‘Tian 00127’ was controlled by one dominant gene to CYR34 and CYR32. The resistance of wheat variety ‘Tian 00127’ to CYR 32 might originate from the parents ‘Baidatou’ or ‘85-173-4’.
Key words inheritance of resistance; seedling stage; stripe rust; Baidatou; Tian 00127
小麦条锈病是甘肃省及中国小麦生产上最主要的病害之一[1],种植抗病品种是防治该病最经济有效的措施[24]。2009年以来,随着以条锈菌CYR34为代表的贵农22致病类群出现频率的不断上升,国内重要抗源材料‘南农92R’‘贵农22’及其衍生系生产品种如‘中梁29号’‘天选43号’‘兰天17号’‘兰天24号’ 等甘肃陇南小麦生产品种田间抗条锈性丧失[57],引致近年来在全国范围内的条锈病流行压力日趋加大,导致2017年小麦条锈病在黄淮海麦区大流行,造成严重的产量和经济损失[89]。近年來的研究发现,甘肃陇南由于条锈菌新菌系的不断出现及以CYR34、CYR32为代表的菌系的联合作用,使得当前陇南种植的小麦品种大都不抗小麦条锈病,特别是苗期抗病品种更是少之又少[10]。进行小麦抗条锈病新材料的评价和利用,对增加该区域抗条锈病基因多样性和丰富度,持续控制条锈病的发生流行具有重要作用。寻找对当前主要流行小种CYR34、CYR32具有良好抗病性的新抗源,是甘肃陇南当前及未来很长一段时期内小麦抗条锈病育种工作中一项需长期解决的问题。
农家小麦品种基因资源优良,是提高小麦抗条锈性的有效途径。明确小麦农家品种及其衍生系材料抗条锈性状况及其遗传基础,可针对性地选育抗病品种,为抗病品种的合理布局奠定基础[11]。农家品种‘白大头’及其衍生系‘天00127’来自天水市农业科学研究所甘谷试验站,近年来在甘肃陇南田间抗病性稳定且表现优异。目前,国内外相关学者利用孟德尔遗传规律,进行了诸多品种抗条锈性遗传分析[1215],但針对农家品种‘白大头’及其衍生系‘天00127’,尚未开展相关研究。基于此,团队开展了供试材料苗期抗条锈性遗传研究,旨在为更好利用该品种提供科学依据。
1 材料与方法
1.1 供试材料
2017年和2018年5月上旬,在甘肃省农业科学院植物保护研究所甘谷试验站,分别以感病材料‘铭贤169’为母本,以‘白大头’和‘天00127’为父本进行杂交、自交和回交,分别获得各杂交组合后代F1、F2和BC1代材料。条锈菌单孢菌系CYR34、CYR32来自甘肃省农业科学院植物保护研究所小麦病害课题组。
1.2 试验方法
苗期抗条锈病鉴定和遗传分析于2019年3月-4月在甘肃省农业科学院植物保护研究所兰州温室进行。相关亲本及杂交F1代、BC1代种子各播种4盆,F2代种子播种24~40盆,BC1代每盆4~10株,亲本和F2代材料每盆20株以上。在小麦生长的1叶1心期,采用抖孢子粉法[7]分别接种供试小种CYR34和CYR32。10℃条件下保湿24 h后置于常规温室生长18 d。感病品种‘铭贤169’充分发病后逐株记载反应型[14]。反应型记载采用0、0;、1、2、3、4 共6级标准进行[16]。为保证准确分析,再加入1+、2-和2+ 3个级别[17]。对实测抗感比率与期望抗感比率的适合性进行χ2检验[18]。
2 结果与分析
2.1 ‘白大头’抗条锈性遗传分析
2.1.1 对CYR34抗条锈性遗传分析
由表1结果看出,亲本‘白大头’对CYR34表现免疫,反应型为0,‘铭贤169’表现高度感病,反应型为4。F1代植株表现免疫或近免疫,反应型为0~0;。F2代群体表现抗感分离。根据双亲‘铭贤169’和‘白大头’及其F1、F2、BC1代植株反应型级别及各级反应型数目,将0~2+型划为抗病类型,3~4型划为感病类型。136株F2代群体中,有32株表现抗病,104株表现感病。经χ2检验,符合由1对隐性基因控制的理论比例(χ2{1∶3}=0.088 2<χ20.05,1=3.84,P=0.05~0.10)。5株BC1代植株全部感病,支持F2代分析结果。表明‘白大头’对CYR34的抗条锈性由1对隐性抗性基因控制。
2.1.2 对CYR32抗条锈性遗传分析
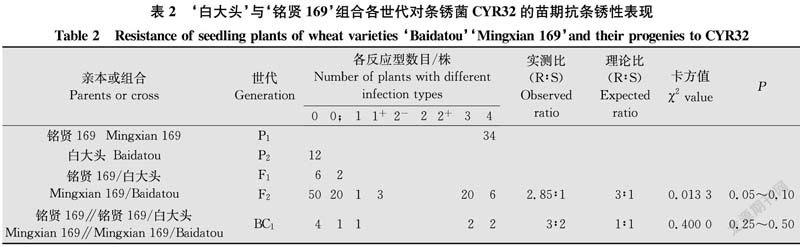
由表2结果看出,亲本‘白大头’对CYR32表现免疫,反应型为0,‘铭贤169’表现高度感病,反应型为4。F1代植株表现免疫或近免疫,反应型为0~0;。F2代群体表现抗感分离。根据双亲‘铭贤169’和‘白大头’及其F1、F2、BC1代植株反应型级别及各级反应型数目,将0~2+型植株划为抗病类型,3~4型划为感病类型。在100株F2代群体中,有74株表现抗病,有26株表现感病。经χ2检验,符合由1对显性基因控制的理论比例(χ2{3∶1}=0.013 3<χ20.05,1=3.84,P=0.05~0.10)。10株BC1代植株,6株表现抗病,4株表现感病,符合由1对显性基因控制的理论比例(χ2{1∶1}=0.400 0,P=0.25~0.50),支持F2代分析结果。表明‘白大头’对CYR32的抗条锈病性由1对显性抗性基因控制。
2.2 ‘天00127’抗条锈性遗传分析
2.2.1 对CYR34抗条锈性遗传分析
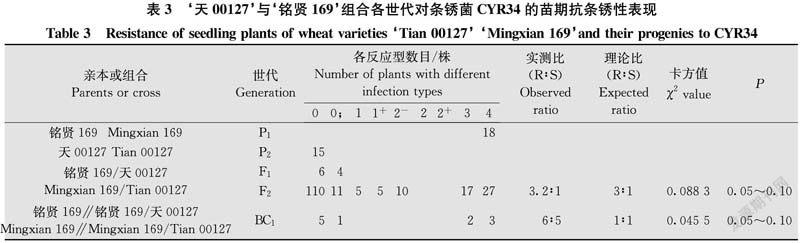
由表3结果看出,亲本‘天00127’对CYR34表现免疫,反应型为0,‘铭贤169’表现高度感病,反应型为4。F1代植株均表现免疫或近免疫,反应型为0~0;。F2代群体表现抗感分离。根据双亲‘铭贤169’和‘天00127’及其F1、F2、BC1代植株反应型级别及各级反应型数目,将0~2+型划为抗病类型,3~4型划为感病类型。185株F2代群体中,有141株表现抗病,有44株表现感病。经χ2检验,符合由1对显性基因控制的理论比例(χ2{3∶1}=0.088 3<χ20.05,1=3.84,P=0.05~0.10)。11株BC1代植株中,抗病株6株,感病株5株,符合由1对隐性基因控制的理论比例(χ2{1∶1}=0.045 5,P=0.05~0.10),支持F2代分析结果。表明‘天00127’对CYR34的抗条锈病性由1对显性抗性基因控制。
2.2.2 对CYR32抗条锈性遗传分析
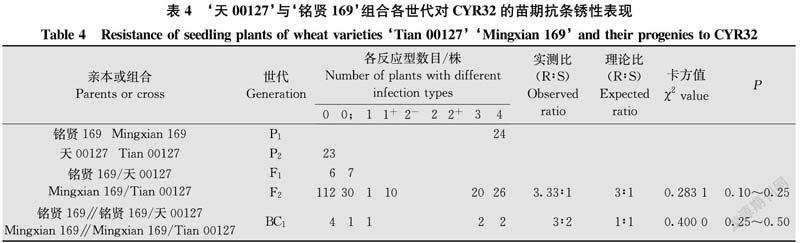
由表4结果看出,亲本‘天00127’对CYR32表现免疫,反应型为0,‘铭贤169’表现高度感病,反应型为4。F1代植株均表现免疫或近免疫,反应型为0~0;。F2代群体表现抗感分离。根据双亲‘铭贤169’和‘天00127’及其F1、F2、BC1代植株反应型级别及各级反应型数目,将0~2+型划为抗病类型,3~4型划为感病类型。在199株F2代群体中,有153株表现抗病,有46株表现感病。经χ2检验,符合由1对显性基因控制的理论比例(χ2{3∶1}=0.283 1<χ20.05,1=3.84,P=0.10~0.25)。10株BC1代植株中,6株表现抗病,4株表现感病,符合由1对显性基因控制的理论比例(χ2{1∶1}=0.400 0,P=0.25~0.50),支持F2代分析结果。表明‘天00127’对CYR32的抗条锈病性由1对显性抗性基因控制。
3 讨论
试验过程中,严格控制试验条件和合理确定抗感分类标准,是小麦抗条锈病性遗传试验结果准确与否的关键所在[19]。本试验在甘肃省农业科学院植物保护研究所兰州温室进行,温室内温湿度条件十分适宜条锈菌生长繁殖,同时试验是在同一批条件下完成,保证了试验结果的准确性[20]。
鉴定及监测结果发现,供试品种‘白大头’苗期对条锈菌主要流行小种CYR34和CYR32均表现免疫,与Ma等[20]研究的其对包括CYR32在内的部分供试条锈菌生理小种表现感病的结果有差异。分析其原因,可能是由于小麦品种的同名异种所致。在品种资源中,同名异种现象极为普遍,只有出自同库、同号材料,才表现出品种自身的特异性、一致性和稳定性特点。
研究结果表明,供试农家品种‘白大头’对CYR34的抗性由1对隐性抗性基因控制,对CYR32的抗性由1对显性抗性基因控制,故推测其含有两个不同的抗条锈病基因。小麦品种‘天00127’为农家品种‘白大头’和高代材料‘C184-3-4-1’及‘85-173-4’的子代材料,其对CYR32的抗性由1对显性基因控制。对其亲本的抗病性分析发现,对CYR32,亲本‘C184-3-4-1’表现感病,‘白大头’和‘85-173-4’表现抗病,故推测‘天00127’对CYR32的抗病基因可能来自‘白大头’或‘85-173-4’。进一步开展‘白大头’及‘天00127’有效抗病基因的标记和利用,将会对进一步充实甘肃陇南抗条锈病基因库和下一步更好地利用奠定基础。
遗传试验中,供试菌系的选择是推断品种抗病基因来源的关键[21]。本试验中,选用致病性较强、且当前为甘肃省主要流行小种的CYR34和CYR32开展苗期抗条锈基因遗传分析,其结果对下一步开展育种和生产应用实践也具有较好的理论指导意义。
参考文献
[1] 李振岐, 曾士迈. 中国小麦锈病[M]. 北京: 中国农业出版社, 2002.
[2] WAN Anmin, ZHAO Zhonghua, CHEN Xianming, et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f.sp. tritici in China in 2002 [J]. Plant Disease, 2004, 88(8): 896904.
[3] MCINTOSH R A, BARINAN H S, PARK R F, et al. Aspects of wheat rust research in Australia [J]. Euphytica, 2001, 119(1/2): 117122.
[4] CHEN Xianming, MARY M, EUQENE A M, et al. Wheat stripe rust epidemics and races of Puccinia striiformis f.sp. tritici in the United States in 2000 [J]. Plant Disease, 2007, 86(1): 3946.
[5] 贾秋珍, 黄瑾, 曹世勤, 等. 感染我国重要小麦抗源材料贵农22的条锈菌新菌系的发现及致病性初步分析[J].甘肃农业科技, 2012(1): 35.
[6] 曹世勤, 贾秋珍, 宋建荣, 等. 甘肃省冬小麦抗条锈菌CYR34育种策略[J].植物遗传资源学报, 2019, 20(5): 11291133.
[7] LIU Taiguo, PENG Yunliang, CHEN Wanquan, et al. First detection of virulence in Puccinia striiformis f.sp. tritici in China to resistance genes Yr24(=Yr26) present in wheat cultivar Chuanmai 42 [J]. Plant Disease, 2010, 94(9): 1163.
[8] 韩德俊, 康振生.中国小麦品种抗条锈病现状及存在问题与对策[J].植物保护, 2018, 44(5): 112.
[9] 黄冲, 姜玉英, 李佩玲, 等. 2017年我国小麦条锈病流行特点及重发原因分析[J].植物保护, 2017, 43(2): 162166.
[10]曹世勤, 王晓明, 贾秋珍, 等. 2003-2013年小麦品种(系)抗条锈性鉴定及评价[J].植物遗传资源学报, 2017, 18(2): 253260.
[11]黄苗苗, 孙振宇, 曹世勤, 等. 223份小麦农家品种田间抗条锈病性评价及抗病基因分子检测[J].植物保护学报, 2017, 45(1): 90100.
[12]姚强, 贺苗苗, 阎佳会, 等. 春小麦品种青春39的抗条锈性遗传分析[J].麦类作物学报, 2014, 34(1): 3942.
[13]李洋, 袁喜麗, 姚强, 等. 欧洲小麦品种Mega抗条锈病基因的遗传分析及分子标记[J].植物病理学报, 2010, 40(1): 5156.
[14]仝淑玫, 蔺瑞明, 何月秋, 等. 小麦抗源Holdfast和Flinor抗条锈病主效、微效基因的遗传分析[J].中国农业科学, 2006, 39(11): 22432249.
[15]曹世勤, 骆惠生, 黄瑾, 等. 冬小麦品种陇鉴9821抗条锈遗传分析[J]. 植物病理学报, 2012, 42(3): 274280.
[16]中华人民共和国农业部. 小麦抗病虫性评价技术规范: 第1 部分 小麦抗条锈病评价技术规范: NT/T1443.1-2007[S]. 北京: 中国农业出版社, 2007.
[17]徐世昌, 张敬原, 赵文生, 等. 小麦京核891-1抗条锈病主效、微效基因的遗传分析[J].中国农业科学, 2001, 34(3): 272276.
[18]何家泌. 植物抗病遗传学[M]. 北京: 中国农业出版社, 1994.
[19]姚占军, 徐世昌, 万安民, 等. 3个小麦条锈菌鉴别寄主的抗性遗传分析[J].植物遗传资源学报, 2006, 7(1): 3943.
[20]MA Dongfang, LI Qiang, TANG Mingshuang, et al. Mapping of gene conferring adult-plant resistance to stripe rust in Chinese wheat landrace Baidatou [J]. Molecular Breeding, 2015, 35: 157162.
[21]徐建龙, 林贻滋, 奚永安. 城特232抗瘟性基因分析研究[J].植物保护学报,1994,21(1):6366.
(责任编辑:杨明丽)
