棉花产量和土壤微生物数量及酶活性对棉田间作系统的响应
2022-02-15崔爱花孙亮庆刘帅白志刚胡启星孙巨龙黄国勤
崔爱花 孙亮庆 刘帅 白志刚 胡启星 孙巨龙 黄国勤



摘要:为探讨红壤旱地上以棉花为主体的间作系统的优势机制,为促进南方旱地农业的绿色、可持续发展提供理论依据,设置大豆、玉米、甘薯3种传统作物分别与棉花间作,以棉花单作为对照,研究棉花产量、土壤微生物数量及酶活性对棉田间作系统的响应。结果表明:(1)间作较单作可通过总铃数的增加来提高籽棉产量,单位面积总铃数增加27.3%~78.7%,2年平均显著增产15.0%~40.4%(P<0.05);(2)各处理的土壤微生物数量均随生育期的推进呈先升后降再升的趋势,间作较单作可显著增加土壤细菌、真菌、放线菌和固氮菌数量,增幅分别为30.7%~64.0%(花铃期)、24.5%~39.0%(蕾期)、23.1%~50.6%(蕾期)和44.1%~94.9%(吐絮期);(3)间作较单作可显著增加土壤中转化酶、脲酶和过氧化氢酶活性,增幅分别为22.8%~73.4%(收获后)、100.0%~250.0%(花铃期)和42.9%~150.0%(吐絮期)。棉花产量、微生物数量及酶活性关系较密切,相互之间存在正向促进作用。棉花甘薯间作可提高土壤中微生物的数量以及酶活性,改善土壤肥力,达到棉花增产的目的,是适宜在红壤旱地上推广应用的较优模式。
关键词:棉花;间作;产量;土壤微生物;酶活性
中图分类号: S344.2;S562.04 文献标志码: A
文章编号:1002-1302(2022)02-0053-06
收稿日期:2021-05-10
基金项目:国家现代农业产业技术体系建设专项(编号:CARS-15-38);江西省农业农村厅科技计划(编号:赣农办字[2017]56、赣农厅办函[2019]76号)。
作者简介:崔爱花(1983—),女,河南新乡人,博士,副研究员,主要从事棉花栽培及生理生态研究工作。E-mail:49856861@qq.com。
通信作者:黄国勤,博士,教授,主要从事作物栽培、耕作制度、农田生态环境等方面的研究。E-mail:hgqjxauhgq@sina.com。
棉花是重要的战略物资,在我国国民经济中的地位尤为重要[1-2],间作在我国应用历史悠久,是一种非常重要的农业生产方式[3],间作优势产生的最主要的生态机制是生态位的分离,这是因为间作中2种作物的地下部与地上部在生态位上发生分离,使时空生态位进行互补扩大,继而使地下的养分资源及地上的光、热资源实现时间的前后分离和空间上的互补扩大,促进资源的高效利用[4-6]。间作以可通过改变土壤微生物数量以及酶活性来影响土壤环境,继而使作物产量及品质发生改变[7-10]。土壤中微生物、酶共同参与土壤的物质循环和能量流动,是评价土壤肥力的重要指标之一[11-13]。本试验探讨土壤微生物及酶活性对红壤旱地棉田间作系统的响应特点,旨在探明棉田间作的优势机制,为解决棉争地矛盾,推进棉花产业的绿色、高效和可持续发展提供参考依据。
1 材料与方法
1.1 试验地概况
于2016—2017年在江西农业大学试验田(115°55′02.040″E,28°46′04.476″N)开展大田试验。试验区光热资源充足,年平均日照时数、日照总辐射分别为1 559.9 h、102.55 kJ/cm2,无霜期约269 d,年均温度及≥10 ℃的活动积温分别为16.5、5 521 ℃,年均降水量为1 658.9 mm。试验初始土壤性状如下:容重1.304 g/cm3,總孔隙度52.98%,毛管空隙度41.55%,有机质含量 29.78 g/kg,pH值4.75,全氮含量1.34 g/kg,碱解氮含量 90.00 mg/kg,全磷含量1.18 g/kg,有效磷含量 76.35 mg/kg,全钾含量55.38 g/kg,速效钾含量107.50 mg/kg。
1.2 试验设计
随机区组设计,以棉花单作(MC)为对照,设3种间作处理,分别是棉花大豆间作(CS)、棉花玉米间作 (CM)和棉花甘薯间作 (CP),共4个处理,每个处理3次重复。小区面积为6.0 m(长)×5.5 m(宽)=33.0 m2。
供试棉花品种为江西省棉花研究所选育的赣棉杂1号;大豆品种选用东北毛豆,玉米品种为甜糯玉米,甘薯品种为南昌农家种,上述作物均从市场购买。
1.3 田间管理
棉花分别于2016年4月25日和2017年4月17日开沟播种,行株距设置为 100 cm×40 cm;大豆和玉米的播种期分别为2016年4月24日和2017年4月16日,种植方式是在2行棉花中间开沟播种,行株距均为100 cm×25 cm。甘薯苗分别于2016年5月11日和2017年4月25日开沟栽种,行株距同大豆和玉米,每种作物的施肥情况如表1所示,其他管理方式同大田一致。
1.4 测定项目和方法
1.4.1 棉花产量和产量性状
棉花产量按实收产量计算;吐絮期,在棉花植株中部位置,每个小区选取30个棉铃,带回室内考种,测出棉花铃质量和衣分;每个小区选取长势一致的10株棉花进行挂牌标记,调查单株成铃数,取平均值。
1.4.2 土壤微生物数量
土壤中的细菌、真菌、放线菌和固氮菌数量采用固体平板培养计数法测定[14]。
1.4.3 土壤酶活性
土壤过氧化氢酶、脲酶及转化酶分别采用高锰酸钾滴定法、靛酚蓝比色法和 3,5-二硝基水杨酸比色法进行测定[15]。
1.5 统计方法
采用Excel 2010对原始数据进行整理,采用SPSS 17.0和OriginPro 8.5软件分别进行数据分析和作图,因2年数据表现趋势较一致,故以2017年的试验结果进行分析说明。
2 结果与分析
2.1 棉田间作对棉花产量及产量构成的影响
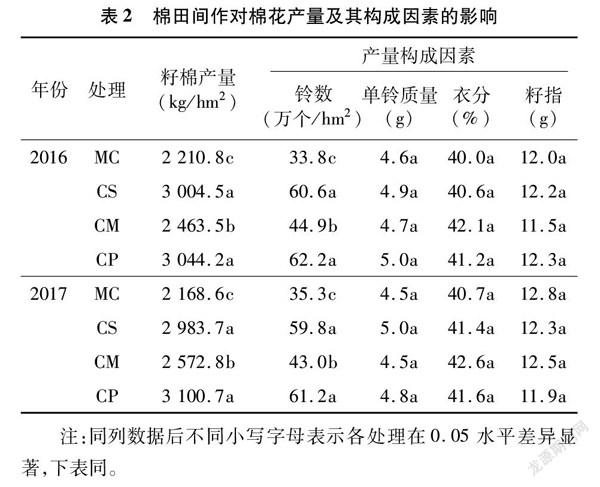
由表2可知,棉田间作可增加棉花产量,2016年CS、CM和CP的籽棉产量分别比MC显著提高35.9%、11.4%和37.7%,2017年CS、CM和CP分别比MC显著提高37.6%、18.6%和43.0%;处理CS和CP之间差异不显著,但两者均显著高于处理CM。各处理单位面积的铃数与籽棉产量表现趋势相似,间作处理铃数较单作年均显著增加27.3%~78.7%;处理CS和CP之间差异不显著,但均显著高于处理CM,与单作相比,间作在增加单铃质量、衣分及籽指方面没有显著作用。因此,棉田间作较单作可有效提高棉花产量,且增产的主要原因是铃数的增加,以处理CS(棉花间作大豆)和处理CP(棉花间作甘薯)的增产优势最为明显。
2.2 棉田间作对土壤微生物数量的影响
由表3可知,各处理的土壤细菌、真菌、放线菌、固氮菌数量均随着生育期的推进呈先升高后降低再升高的趋势。
在土壤微生物的主要组成成分中,细菌生物量占比最大,它可以有效分解各种有机物,是衡量土壤肥力的重要指標之一[16-17]。细菌数量的增加可提高土壤微生物活性,能够促进植物生长,增加产量。处理MC和CP的土壤细菌数最高值出现在蕾期,CS和CM的最高值在花铃期。棉花播种前、蕾期和花铃期处理CS、CM和CP的细菌数量均显著高于MC,增幅分别为36.4%~65.6%、23.8%~31.2%和30.7%~64.0%;花铃期至吐絮期下降幅度较大,吐絮期处理CS和CP分别较处理MC显著增加34.5%和60.3%,处理CM与MC之间差异不显著;吐絮期以后各处理土壤细菌数量有所上升,棉花收获后,处理CS较处理MC显著高33.6%,处理CM和CP与处理MC之间差异均不显著。以上结果表明,间作较单作可有效提高土壤细菌含量,其中处理CS(棉花间作大豆)表现最优。
处理MC、CS和CM的真菌数量最高值均出现在蕾期,处理CP的最高值在花铃期。播种前,各处理的土壤真菌含量差异不显著,随后明显增加,至蕾期,处理CS、CM和CP分别较处理MC显著增加37.3%、24.5%和39.0%,处理CP显著高于处理CM,但与CS之间差异不显著。花铃期,处理CS和CP较处理MC分别显著增加22.8%和45.5%;处理CM与处理MC之间差异不显著;花铃期至吐絮期,各处理真菌数量急剧下降,吐絮期,间作处理CS、CP分别比MC显著低41.8%、29.1%;吐絮期以后各处理土壤真菌数量有所上升,棉花收获后,各处理之间无显著差异。以上结查表明,棉田间作较单作可有效提高棉花蕾期和花铃期的土壤真菌数量,处理CP优势最明显。
处理CS、CM和CP的放线菌数量最高值均出现在蕾期,处理MC的最高值在花铃期。各处理的放线菌数量除在花铃期无显著差异外,其余各时期均存在一定的差异。棉花播种前和收获后均表现为处理CS、CM和CP之间差异不显著,不同的是,播种前仅处理CP较MC显著高65.6%,收获后处理CS和CM较处理MC显著高35.8%和25.4%。蕾期,处理CS、CM和CP分别较处理MC显著增加50.6%、30.5%和23.1%;处理CS显著高于处理CP,但与处理CM之间差异不显著。吐絮期,各处理的土壤放线菌数量降到最低,处理CP较处理MC显著高80.0%,处理CS、CM与处理MC之间差异均不显著。综合分析认为,处理CP在增加土壤放线菌数量方面优势明显。
各处理的固氮菌数量最高值均出现在蕾期。棉花播种前和花铃期各处理表现的趋势相同,处理CS和CP均显著高于处理MC,而处理CM与处理MC之间差异不显著。不同点是播种前处理CS和CP均显著高于处理CM,而花铃期则是3个间作处理之间差异均不显著;吐絮期和收获后均表现为间作处理CS、CM和CP之间差异不显著,不同的是吐絮期,处理CS、CM和CP均显著高于处理MC,增幅为44.1%~94.9%;而收获后,仅处理CP显著高于MC,增幅为58.3%。由以上结果可知,棉田间作可增加土壤固氮菌的数量,以处理CP表现最好。
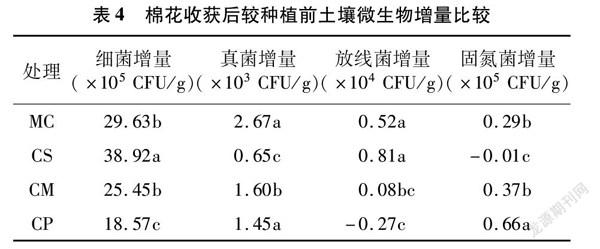
将各处理棉花收获后较种植前土壤微生物数量的增量列于表4,可知除CP处理放线菌和CS处固氮菌的增量为负值外,各处理的土壤微生物数量均有不同程度的增加,与单作相比,CS对细菌、放线菌生物量增加有明显优势;CP对固氮菌生物量增加有明显优势。综合来看,CP优势明显。
2.3 棉田间作对土壤酶活性的影响
土壤酶活性的大小受植物种类、养分含量、土壤质地等多种因素的影响,常作为土壤肥力的主要衡量标准之一[6]。
2.3.1 棉田间作对土壤转化酶活性的影响
转化酶常用于衡量土壤肥力水平和熟化程度,参与土壤中碳循环过程,对土壤中易溶性营养物质的增加起着重要作用。由图1可知,处理MC的土壤转化酶活性随着生育进程的推进呈先升高后降低的趋势,最高值出现在吐絮期,处理CS、CM和CP均呈先升后降再升的趋势,最高值出现在花铃期或收获后。除花铃期外,其余各时期均有间作处理与单作处理存在显著性差异。播种前,间作各处理均显著高于MC,增幅为28.7%~72.3%。蕾期,处理CS比处理MC显著高44.4%,而处理CM和CP均与处理MC之间差异不显著。吐絮期,处理CS、CM和CP均显著低于处理MC,降幅为26.3%~40.4%。收获后,间作处理均显著高于处理MC,增幅为 22.8%~73.4%。
2.3.2 棉田间作对土壤脲酶活性的影响
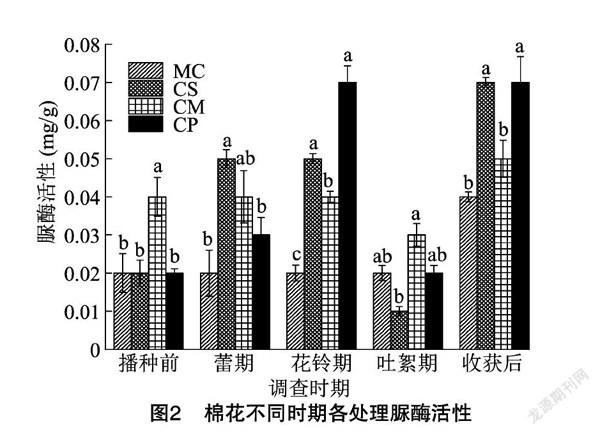
由图2可知,处理MC土壤脲酶活性播种前至吐絮期均无明显变化,收获后达到最高值;处理CS和CP土壤脲酶活性均呈先升后降再升高的趋势,不同的是,处理CS在蕾期至花铃期土壤脲酶活性无变化,而处理CP在花铃期和收获后差异不明显;处理CM土壤脲酶活性,播种前至花铃期无明显差异,花铃期至吐絮期下降而后升高。播种前,处理CM土壤脲酶活性最高,比处理MC、CS和CP均高100%。蕾期,处理CS比处理MC高150.0%,两者差异显著(P<0.05),而处理CM和CP均与处理MC之间差异不显著。花铃期,处理CP土壤脲酶活性最高,其次是处理CS和CM,三者分别比处理MC高250.0%、150.0%和100.0%,差异显著(P<0.05)。吐絮期,处理CS、CM和CP均与处理MC之间无显著差异,处理CM显著高于处理CS。收获后,处理CS和CP均比处理MC高75.0%,差异显著(P<0.05),而处理CM与处理MC之间差异不显著。
2.3.3 棉田间作对土壤过氧化氢酶活性的影响
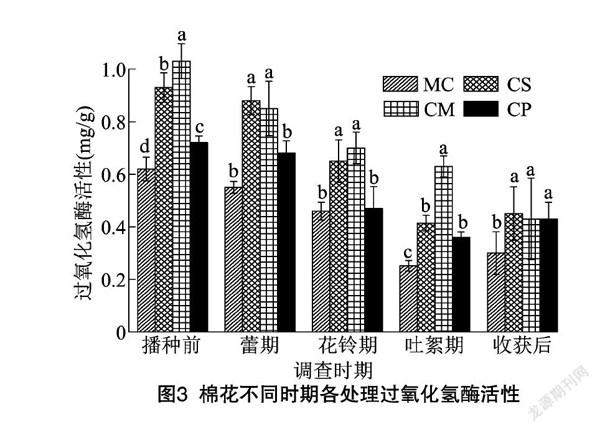
由图3可以看出,處理MC、CS和CP的土壤过氧化氢酶活性均随着生育期的推进呈先降低后升高的趋势,拐点均在吐絮期;而处理CM呈逐渐降低的趋势。播种前,处理CS、CM和CP分别显著比处理MC高50.0%、66.1%和16.1%;蕾期,处理CS、CM之间差异不显著,分别显著比处理MC高60.0%和54.5%,处理CP与处理MC之间差异不显著;花铃期与蕾期表现一致,亦是处理CS和CM差异不显著,分别显著比处理MC高41.3%和52.2%,而处理CP与MC之间差异不显著;吐絮期,处理CM最高,处理CS和CP次之,分别显著比处理MC高150.0%、64.3%和42.9%;收获后,处理CS、CM和CP之间差异不显著,分别显著比处理MC高50.0%、43.3%和43.3%。
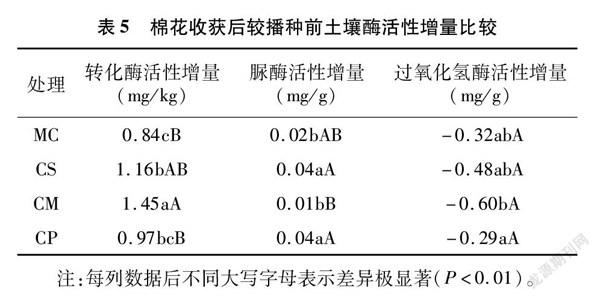
将棉花收获后较播种前的土壤酶活性增量列于表5,处理CS、CM在促进转化酶活性增加方面优势明显;处理CS和CP较MC可有效增加脲酶活性;各处理的土壤过氧化氢酶活性增量均为负值,处理CP优势较明显。
2.4 土壤微生物数量、酶活性与棉花产量的相关性
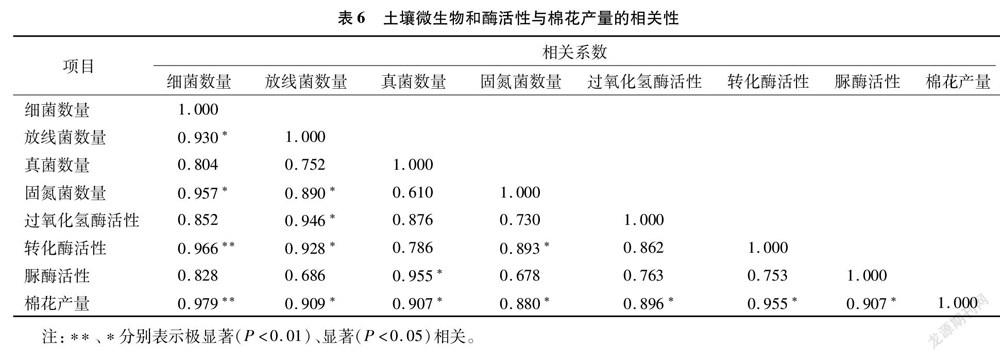
将土壤微生物中的细菌、放线菌、真菌及固氮菌数量,过氧化氢酶、脲酶、转化酶活性以及棉花产量等8个指标之间的相关关系进行分析。由表6可知,各指标之间均存在正相关关系,棉花产量除与细菌数量达到极显著相关外,与其余各指标之间的相关性均达到显著水平,说明土壤微生物数量及酶活性直接影响到棉花产量。另外,细菌数量分别与放线菌、固氮菌数量存在显著相关关系,与转化酶活性的相关性达到极显著水平;固氮菌、放线菌数量及转化酶活性彼此之间均呈显著相关;真菌数量与脲酶活性、放线菌数量与过氧化氢酶和转化酶活性均达到显著相关。
3 讨论
孙建等认为,土壤微生物在土壤生态系统中具有重要作用,与农田生态系统关系密切[12]。众多学者开展了有关土壤微生物对间作系统的响应研究,得出的结论是,通过间作种植可使土壤系统中的微生物数量及多样性均有不同程度的增加。如沈雪峰等研究表明,甘蔗与花生间作能显著增加土壤中的细菌、放线菌及真菌的数量[18];宋亚娜等开展了小麦‖蚕豆、玉米‖蚕豆和小麦‖玉米间作效应研究,证实了间作可提高作物根际微生物的多样性[19];张向前等一致认为,间作较单作显著增加了土壤中的细菌、真菌、固氮菌的数量[8,20]。但张亮亮等在研究结论中指出,与棉花单作相比,南疆的枣‖棉间作在促进土壤微生物数量增加方面的作用不显著[21]。本研究中的土壤细菌和真菌数量在棉田间作条件下均得到了有效提高;棉花与大豆间作处理的土壤放线菌数量明显增加,棉花与甘薯间作处理在增加土壤中的固氮菌数量方面有优势。
李成芳等在研究中指出,土壤酶活性和农业措施及土壤肥力存在显著相关关系,可反映土壤中多种生化过程强度[22]。刘均霞等提出,间作作物根际的土壤磷酸酶和脲酶活性显著高于任一单作[7];覃娟等研究认为,甘蓝间作水萝卜处理较单作促进了不同耕层土壤中过氧化氢酶、蔗糖酶和脲酶活性的增加,差异达到了显著或极显著水平[23];但谢利等在棉花‖孜然间作模式对土壤微生物数量及酶活性的影响的研究中得出,间作较单作使土壤中的过氧化氢酶、蔗糖酶及脲酶的活性呈不同程度的降低,差异达到显著水平[24]。本研究中棉田间作模式较棉花单作可有效促进土壤中3种酶活性的增加。
产量与土壤生态系统中的微生物数量及酶活性密切相关,存在正的相关关系,相关性达到极显著或显著水平[8,25-27]。符冠富等研究指出,土壤酶活性在水稻的灌浆中后期与产量以及干物质量存在显著相关关系[28];马冬云等得出了小麦产量、根际土壤系统中的酶活性和微生物数量存在显著正相关关系的结论[29]。本研究与上述研究结论一致,土壤微生物数量、棉花产量及酶活性彼此之间均存在正的相关关系,棉花产量除与细菌数量达到极显著相关外,与其余各指标之间的相关性均达到显著水平。
4 结论
在红壤旱地棉田实施间作均能不同程度地增加单位面积棉花成铃数和籽棉产量;提高棉田土壤中的真菌、细菌、放线菌和固氮菌数量及微生物总数,可有效增加土壤过氧化氢酶、脲酶和转化酶活性,以棉花间作甘薯模式较好;棉花产量与微生物数量及酶活性三者之间存在正相关关系。因此,大力发展以棉花为主体作物的间作,不仅可以缓解粮棉争地矛盾,且可以提高资源利用效率,达到增产增收的目的,促进旱地农业的绿色、高效和可持续发展。
参考文献:
[1]王子胜,徐 敏,张国伟,等. 施氮量和种植密度对东北特早熟棉区棉花生物量和氮素累积的影响[J]. 应用生态学报,2011,22(12):3243-3251.
[2]刘 洋,李亚雄,李 斌,等. 我国棉花加工机械关键技术的专利分析[J]. 新疆农机化,2015(3):41-43.
[3]刘巽浩,牟国正. 中国耕作制度[M]. 北京:中国农业出版社,1993:7-15.
[4]肖焱波,李 隆,张福锁. 小麦/蚕豆间作体系中的种间相互作用及氮转移研究[J]. 中国农业科学,2005,38(5):965-973.
[5]肖焱波,李 隆,张福锁. 根瘤菌菌株NM353对小麦/蚕豆间作体系中作物生长及养分吸收的影响[J]. 植物营养与肥料学报,2006,12(1):89-96.
[6]李 隆. 间作作物种间促进与竞争作用研究[D]. 北京:中国农业大学,1999:100-101.
[7]刘均霞,陆引罡,远红伟,等. 玉米、大豆间作对根际土壤微生物数量和酶活性的影响[J]. 贵州农业科学,2007,35(2):60-61.
[8]张向前,黄国勤,卞新民,等. 间作对玉米品质、产量及土壤微生物数量和酶活性的影响[J]. 生态学报,2012,32(22):7082-7090.
[9]张海春,张 浩,胡晓辉. 不同间作模式对温室连作番茄产量、土壤微生物和酶活性的影响[J]. 西北农业学报,2016,25(8):1218-1223.
[10]徐 双,柳新伟,崔德杰,等. 不同施肥处理对滨海盐碱地棉花生长和土壤微生物及酶活性的影响[J]. 水土保持学报,2015,29(6):316-320.
[11]苗 琳,王 立,黄高宝,等. 保护性耕作对旱地麦田土壤酶活性的影响[J]. 干旱地区农业研究,2009,27(1):6-11.
[12]孙 建,刘 苗,李立军,等. 免耕与留茬对土壤微生物量C、N及酶活性的影响[J]. 生态学报,2009,29(10):5509-5515.
[13]姜 莉,陈源泉,隋 鹏,等. 不同间作形式对玉米根际土壤酶活性的影响[J]. 中国农学通报,2010,26(9):326-330.
[14]中国科学院南京土壤所微生物室. 土壤微生物研究法[M]. 北京:科学出版社,1985:12-36.
[15]关松荫. 土壤酶及其研究法[M]. 北京:农业出版社,1986:274-325.
[16]沈其荣,褚贵新,曹金留,等. 从氮素营养的角度分析旱作水稻与花生间作系统的产量优势[J]. 中国农业科学,2004,37(8):1177-1182.
[17]Zhang X Q,Huang G Q,Bian X M,et al. Effects of nitrogen fertilization and root interaction on the agronomic traits of intercropped maize,and the quantity of microorganisms and activity of enzymes in the rhizosphere[J]. Plant and Soil,2013,368(1):407-417.
[18]沈雪峰,方 越,董朝霞,等. 甘蔗/花生間作对土壤微生物和土壤酶活性的影响[J]. 作物杂志,2014(5):55-58.
[19]宋亚娜,Marschner P,张福锁,等. 小麦/蚕豆、玉米/蚕豆和小麦/玉米间作对根际细菌群落结构的影响[J]. 生态学报,2006,26(7):2268-2274.
[20]吴凤枝,周新刚. 不同作物间作对黄瓜病害及土壤微生物群落多样性的影响[J]. 土壤学报,2009,46(5):899-906.
[21]张亮亮,罗 明,韩 剑,等. 南疆枣树-棉花间作对土壤微生物区系及代谢熵的影响[J]. 棉花学报,2016,28(5):493-503.
[22]李成芳,曹凑贵,徐拥华,等. 稻鸭与稻鱼生态系统土壤微生物量N和土壤酶活性动态[J]. 生态学报,2008,28(8):3905-3912.
[23]覃 娟,李 剑,刘 霞,等. 甘蓝水萝卜间作对土壤理化性状及酶活性影响[J]. 北方园艺,2010(7):20-22.
[24]谢 利,王燕芳,马 超,等. 棉花-孜然间作模式对土壤微生物数量及酶活性的影响[J]. 江苏农业科学,2015,43(10):103-104.
[25]张继光,秦江涛,要文倩,等. 长期施肥对红壤旱地土壤活性有机碳和酶活性的影响[J]. 土壤,2010,42(3):364-371.
[26]崔爱花,黄国勤. 间作对棉花产量、土壤微生物数量及酶活性的影响[J]. 江西农业学报,2021,33(4):22-26.
[27]马洪波,杨 苏,李传哲,等. 不同肥料和生物菌剂对重茬甘薯产量及土壤质量的影响[J]. 江苏农业科学,2019,47(24):47-49,57.
[28]符冠富,王丹英,徐春梅,等. 稻田冬季保护性耕作条件下的土壤酶活性与水稻成熟期叶片衰老和籽粒产量之间的关系[J]. 中国水稻科学,2009,23(1):43-50.
[29]马冬云,郭天财,宋 晓,等. 尿素施用量对小麦根际土壤微生物数量及土壤酶活性的影响[J]. 生态学报,2007,27(12):5222-5228.
