不同来源地茶树种质资源表型性状遗传多样性分析
2021-12-08冯花,王飞权,陈荣冰,张渤,庄晓芸,刘梦娜,曾紫青
冯花,王飞权,陈荣冰,张渤,庄晓芸,刘梦娜,曾紫青


摘 要:為了对不同来源地茶树种质资源进行鉴定评价和遗传多样性分析,以来自福建、广东、台湾的72份茶树种质作为研究对象,对其27项表型性状进行观测与分析。结果表明:72份茶树种质资源遗传变异性丰富,平均遗传多样性指数(H′)为1.30,其中数量性状(1.82)大于质量性状(0.94),以梗粗的最大(2.15);数量性状的平均变异系数(CV)为17.86%,以百芽重的最大(29.21%),其次是发芽密度(23.46%)。相关性分析发现多个数量性状间的关系复杂,有22对性状的相关性达极显著水平(P?0.01),8对达显著水平(P?0.05);聚类分析结果显示,72份茶树种质在遗传距离为16时被划分为4个类群,各类群间的主要性状差异显著或极显著,且形态特征和进化类型各异;主成分分析表明,前10个主成分的特征值大于1,代表了27项表型性状76.04%的信息;根据主成分综合得分大小,筛选出综合得分前5的茶树种质可在茶叶新产品开发、茶树育种等方面加以利用。
关键词:茶树种质资源;表型性状;遗传多样性
中图分类号:S571.1 文献标识码:A
Genetic Diversity Analysis of Phenotypic Characters of Tea Germplasm Resources from Different Origins
FENG Hua, WANG Feiquan*, CHEN Rongbing*, ZHANG Bo, ZHUANG Xiaoyun, LIU Mengna, ZENG Ziqing
College of Tea and Food Science, Wuyi University / Tea Science Research Institute of Wuyi University, Wuyishan, Fujian 354300, China
Abstract: In order to evaluate and analyze the genetic diversity of the tea germplasm resources from different regions, 72 kinds of germplasm resources from Fujian, Guangdong and Taiwan were selected as the research objects, and 27 phenotypic characters were observed and analysed. The 72 tea germplasms were rich in hereditary variability. The average genetic diversity index (H′) was 1.30 while the index of stem thick of tea was the biggest (2.15), and among them the quantitative character (1.82) was higher than that of the qualitative character (0.94). The average coefficient of variation (CV) of quantitative character was 17.86% while the biggest was the hundred–bud weight (29.21%), followed by the index of germination density (23.46%). Besides, the correlation analysis found that the relationship between multiple quantitative traits was complex. The correlation of 22 pairs of characters reached extremely significant level (P?0.01), and that for 8 pairs of characters reached significant level (P?0.05). Cluster analysis showed that the 72 germplasm were divided into four groups when the genetic distance was 16, the main characters of each group were significantly different, and the various morphological characters and evolutionary types were different. Principal component analysis showed that the characteristic number of the first 10 principal components were greater than 1, which representing 76.04% information of 27 phenotypic characters. The top five germplasm were selected, which could be used in the new products development, tea breeding based on the comprehensive score of the principal components. This study would provide some references for the development and utilization of the germplasm resources of tea and the breeding of new cultivars.
Keywords: tea germplasm resources; phenotypic character; genetic diversity
DOI: 10.3969/j.issn.1000-2561.2021.10.003
福建、广东和台湾是中国乌龙茶的传统主产区[1]。在这些产区,分布着丰富的、适制乌龙茶的茶树种质资源,如‘黄旦‘福建水仙等品种资源,‘武夷名丛‘凤凰单丛等地方种质资源,‘黄观音‘瑞茗等创新种质[2-8],为乌龙茶新产品开发、种质创新及育种工作奠定了物质基础。茶树表型性状包括树型、树姿及新梢、叶片、花器官性状等多项指标[9],利用表型性状鉴定评价茶树种质资源是种质资源研究的常用方法[10]。研究表明,芽叶色泽、叶质、百芽重、发芽密度、树型、树姿等表型性状与茶树次生代谢、茶叶品质、产量及遗传进化密切相关[11-18]。因此,基于表型性状对来自福建、广东、台湾的茶树种质资源进行鉴定评价,筛选高产、优质茶树种质,对促进乌龙茶新产品开发、新品种选育及产业可持续发展具有重要意义。
近年来,基于生化成分[19-20]、代谢[21]、农艺性状[22]、解剖结构[23]、矿质元素[24]及分子标记技术[25-28],对福建武夷山、闽南、广东等地方茶树种质资源的鉴定评价,虽有诸多报道,但因研究范围较窄,尚不能全面揭示适制乌龙茶茶树种质资源的遗传多样性。而对于不同来源地适制乌龙茶茶树种质资源的鉴定评价,仅有少数报道从分子水平上揭示了来自于福建、广东、台湾茶树种质资源的遗传多样性与亲缘关系[29-31],基于表型性状的鉴定评价与遗传多样性分析尚未见报道。研究认为,表型多样性是植物遗传物质多样性的直接体现,基于表型性状分析种质资源遗传多样性是最直观、最基础的研究方法,也是植物优良品种选育的基础[32-33]。茶树作为典型的叶用植物,其表型性状是选择茶树育种材料的重点[34],基于表型性状开展资源的鉴定评价与遗传多样性分析,已在陕西[10,35-36]、浙江[37]、四川[34]、云南[38-40]、贵州[41]、江苏[42]、河南[43]、广西[44]、安徽[45]、江西[46]等地方茶树种质资源的研究中广泛应用,并取得显著成效。因此,本试验以武夷学院茶树种质资源圃保存的来自福建福安、安溪、平和、建瓯、建阳、武夷山,以及广东潮安和台湾的72份茶树种质为研究对象,在同一生境和田管条件下,对其27项表型性状进行观测、分析与综合评价,旨在揭示不同来源地茶树种质资源的遗传多样性与亲缘关系,筛选综合性状优良的茶树种质,为茶树种质资源的开发利用、新品种选育提供参考。
1 材料与方法
1.1 材料
供试材料为保存在武夷学院茶树种质资源圃的72份茶树种质,其来源地和综合得分信息见表1。资源圃位于福建省武夷山市武夷学院四桥旁(27°44'20″ N,117°59′51″ E),属中亚热带季风湿润气候区,光照充足(年平均日照时数1629.5 h),雨量丰沛(年平均降雨量1926.9 mm),温湿度适宜(年平均气温18.3 ℃,10 ℃以上的活动积温在5000 ℃以上,相对湿度在80%左右),适合茶树生长[47]。圃内每个小区保存1份茶树种质,小区长、宽为20 m×1.8 m,双行单株条列式种植,小行距×株距为0.4 m×0.3 m,每份种质种植150株,其中顶端4株为自然生长,其余正常修剪。资源圃地势平缓,立地条件和栽培管理措施基本一致,即年施基肥1次、追肥2次,人工除去杂草,每年一耕三锄,7月和11月各修剪1次,并根据病虫害发生情况及时防治。
1.2 方法
在同一生境和田间管理条件下,按照茶树种质资源性状描述与鉴定方法[48-50]及乌龙茶制造对鲜叶原料形态特征要求[17],于2017、2018年春季,对茶树种质的树型、树姿、芽叶色泽、芽叶茸毛、发芽密度、一芽三叶长、一芽三叶百芽重(以下简称“百芽重”)、梗粗、节间长进行观测,其中节间长和梗粗分别为驻芽小开面第一至第四叶间嫩茎的长度和嫩茎的平均粗度;同年秋季,对叶片着生状态、叶长、叶宽、叶形、测脉对数、叶色、叶面隆起性、叶身、叶质、叶齿锐度、叶齿密度、叶齿深度、叶基、叶尖、叶缘及花冠直径进行观测。其中,对树姿等16项质量性状进行赋值化处理[49],以便统计分析(表2)。
1.3 数据处理
对表型数据,采用Microsoft Excel 2010软件进行基本统计及频率分布、变异系数、遗传多样性指数的分析,采用IBM SPSS 20.0软件进行相关性分析(Person)、系统聚类、主成分分析与综合评价。质量性状的级别按照表2进行划分,各级别频率为相应级別种质数量占总数的百分比;遗传多样性指数(H′)采用Shannon- Weave指数法进行计算[51];主成分分析采用因子过程的主成分分析法,提取特征值大于1.0的因子作为主成分;聚类分析采用离差平方和法(Wards method),遗传距离为欧氏距离[52],采用One-Way ANOVA法和最小显著差异法(LSD)对各类群的表型性状进行方差分析和多重比较。
2 结果与分析
2.1 茶树种质资源表型变异与多样性分析
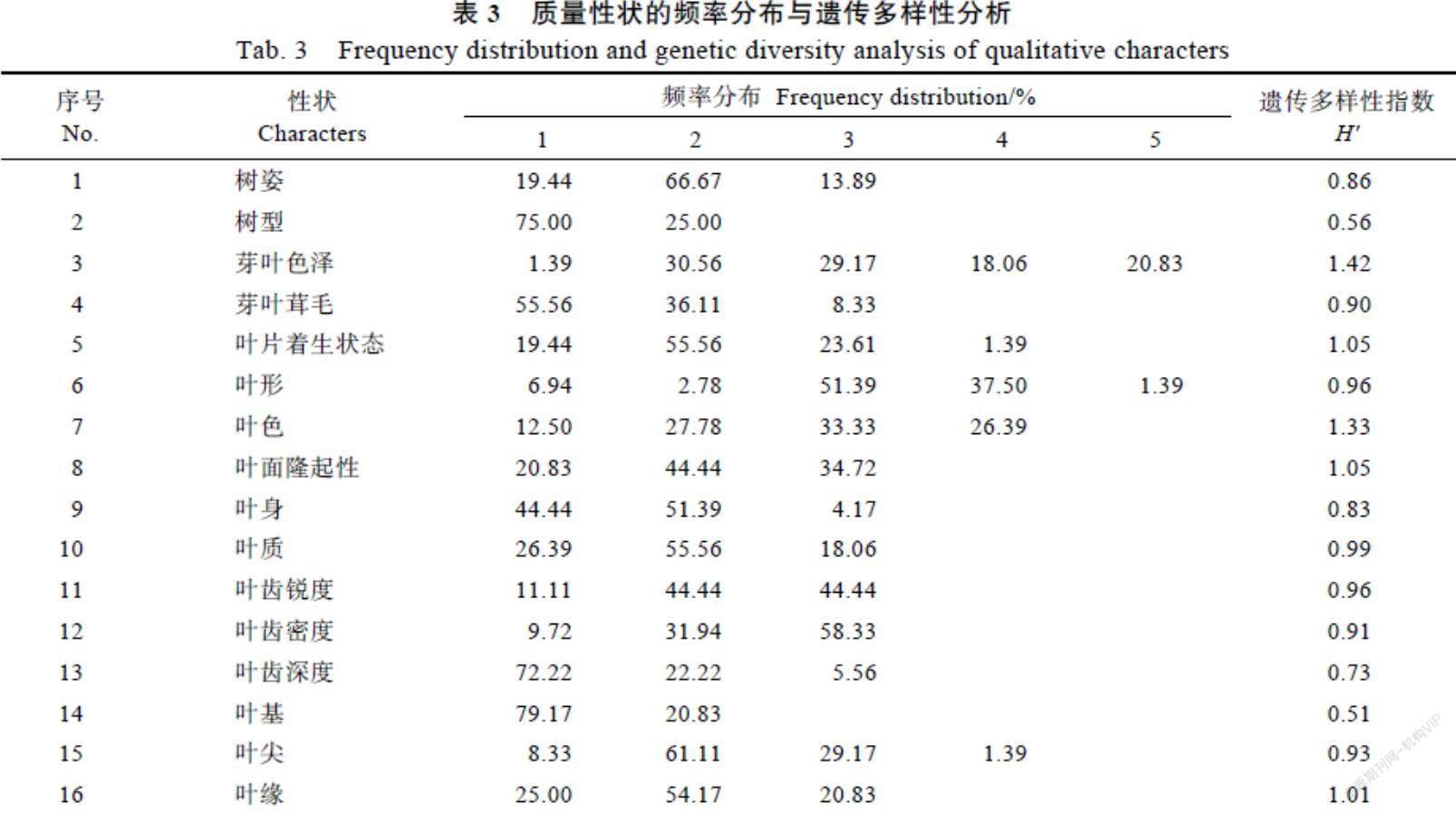
2.1.1 质量性状的频率分布与多样性分析 72份茶树种质资源的质量性状频率分布结果显示
(表3),其树姿、树型和叶片着生状态以半开张、灌木型和稍上斜为主,芽叶色泽以黄绿色、浅绿色或紫绿色为主,芽叶茸毛以少或中为主,叶形、叶质、叶基、叶尖和叶缘分别以椭圆形或长椭圆形、中、楔形、渐尖和微波为主,叶色多浅绿色或绿色,叶面多微隆起或隆起,叶身多内折或平,叶齿主要表现为密、中或钝、浅。16项质量性状的遗传多样性指数(H')在0.51~1.42之间,平均为0.94,其中芽叶色泽、叶片着生状态、叶色、叶面隆起性和叶缘的较大,分别为1.42、1.05、1.33、1.05和1.01,树型和叶基的较小,分别为0.56、0.51,表明72份茶树种质资源芽叶色泽等5个性状的变异类型丰富、分布均匀,可作为茶树品种选育和遗传改良的评价指标。
2.1.2 数量性状的变异与多样性分析 72份茶树种质资源的数量性状的统计分析结果显示(表4),11项数量性状的遗传多样性指数(H')在1.35~2.15之间,平均为1.82,明显大于质量性状,说明数量性状的遗传改良潜力大于质量性状。其中,梗粗、节间长的H'最高,分别为2.15和2.11,一芽三叶长、发芽密度、花冠直径和叶长/叶宽次之,分别为1.98、1.94、1.93和1.89,叶宽最低(1.35),表明72份茶树种质资源在梗粗、节间长、一芽三叶长、发芽密度、百芽重和叶长/叶宽等性状上具有较大的遗传改良潜力。
种质资源表型性状的变异系数代表了某性状变量离散程度的绝对值[53]。结果显示,11项数量性狀的变异系数(CV)在12.46%~29.21%之间,平均为17.86%,其中百芽重最大(29.21%),变化范围在40.90(WL42)~158.50(WL06)g之间;发芽密度、一芽三叶长、梗粗/节间长次之,CV分
别为23.46%、19.47%、18.77%,变化范围分别为40.00(WL52)~128.00(WL45)个/0.11 m2、3.96(WL67)~10.37(WL11)cm、0.12(WL11)~0.37(WL45)mm/cm;叶长/叶宽的CV最小(12.46%),变化范围在1.61(WL44)~3.37(WL59)之间,表明72份茶树种质资源在数量性状上均发生较大幅度的变异,个体间差异较大,其中百芽重、发芽密度、一芽三叶长、梗粗/节间长具有较大的研究价值和选择潜力。
2.2 茶树种质资源数量性状相关性分析
数量性状相关性分析结果显示(表5),16对性状间呈极显著(P?0.01)正相关、4对显著(P?0.05)正相关、6对极显著负相关、4对显著负相关。其中,百芽重与叶长、叶宽、一芽三叶长、节间长、梗粗极显著正相关,与发芽密度、叶长/叶宽、梗粗/节间长显著或极显著负相关;
发芽密度与梗粗/节间长显著正相关,与叶宽显著负相关;一芽三叶长与叶长、叶宽、百芽重、节间长极显著正相关,与梗粗/节间长极显著负相关;叶宽与叶长、百芽重、一芽三叶长、侧脉对数、节间长、梗粗显著或极显著正相关,与叶长/叶宽、发芽密度显著或极显著负相关;梗粗与叶宽、百芽重、节间长、梗粗/节间长显著或极显著正相关,与叶长/叶宽显著负相关;梗粗/节间长与发芽密度、梗粗显著或极显著正相关,与百芽重、一芽三叶长、节间长显著或极显著负相关;叶长/叶宽与叶宽、百芽重、梗粗显著或极显著负相关;花冠直径与叶长、叶宽、百芽重、一芽三叶长极显著正相关,与叶长/叶宽、发芽密度显著负相关。结果表明,72份茶树种质资源多个数量性状间存在着互相促进、互相制约的复杂关系。
2.3 茶树种质资源的聚类分析
从图1可见,基于27项表型性状将72份茶树种质划分为4个类群。其中,第Ⅰ类群的种质数量占总数的9.72%,包括WL01、WL05、WL06等7份茶树种质;第Ⅱ类群的种质数量占总数的52.78%,其中第Ⅱ-ⅰ亚群有WL02、WL07、WL09等31份茶树种质,第Ⅱ-ⅱ亚群有WL28、WL30、WL39等7份茶树种质;第Ⅲ类群的种质数量占总数的30.56%,其中第Ⅲ-ⅰ亚群有WL10、WL14、WL16等10份茶树种质,第Ⅲ-ⅱ亚群有WL04、WL08、WL17等12份茶树种质;第Ⅳ类群的种质数量占总数的6.94%,包括WL63、WL64、WL65等5份茶树种质。结果表明,来自福建、台湾的茶树种质资源均聚在前3个类群,说明两个来源地之间具有较近的亲缘关系,而来自广东的种质主要聚在第4类群,与其他来源地的亲缘关系较远。
方差分析结果显示(表6),4个类群间在树姿、叶片着生状态、叶面隆起性、叶齿锐度与密度、叶缘和发芽密度上差异不显著,在叶齿深度、芽叶茸毛和梗粗上差异显著(P?0.05),在树型、叶形、叶长、叶宽、叶长/叶宽、侧脉对数、叶色、叶身、叶质、叶基、叶尖、芽叶色泽、百芽重、一芽三叶长、节间长、梗粗/节间长和花冠直径上差异极显著(P?0.01)。其中,第Ⅰ类群的叶形、叶宽、叶长/叶宽、叶基、叶尖、百芽重、一芽三叶长与其他类群差异显著,表现为近圆或卵圆的叶形、叶宽、叶长/叶宽值小,近圆形叶基、叶尖钝尖、百芽重大、一芽三叶长、梗粗长、发芽密度小、花冠直径大的特点;第Ⅱ类群的芽叶茸毛与其他类群差异显著,表现为芽叶茸毛多、其他性状多居中、花冠直径小的特点;第Ⅲ类群的树型、叶长、侧脉对数、叶齿深度、梗粗与其他类群差异显著,表现为灌木树型、叶长小、侧脉对数少、叶齿浅、梗细、花冠直径小的特点;第Ⅳ类群的树型、侧脉对数、叶色、叶质与其他类群差异显著,表现为小乔木树型、叶片侧脉对数多、叶色浅、叶质柔软、百芽重小、发芽密度大和花冠直径大的特点。
2.4 茶树种质资源表型性状主成分分析与综合评价
2.4.1 主成分分析 27项表型性状被综合为10个主成分,其累积贡献率为76.04%,包含了表型性状的大部分信息,可以进行综合评价(表7)。
其中,第1主成分方差贡献率为16.64%,主要反映了叶形、叶宽、叶长/叶宽、叶基和叶尖的信息;第2主成分方差贡献率为11.43%,主要反映了百芽重、一芽三叶长、节间长、梗粗/节间长的信息;第3主成分方差贡献率为10.89%,主要反映了叶长、叶宽、侧脉对数和花冠直径的信息;第4主成分方差贡献率为7.95%,主要反映了叶齿锐度和叶齿深度的信息;第5主成分方差贡献率为6.42%,主要反映了树型、芽叶色泽、梗粗的信息;第6主成分方差贡献率为5.69%,主要反映了叶色和叶质的信息;第7主成分方差贡献率为4.88%,主要反映了叶面隆起性和叶身的信息;第8、第9主成分方差贡献率分别为4.30%、4.13%,分别反映了芽叶茸毛、树姿与叶齿密度的信息;第10主成分方差贡献率为3.72%,主要反映了发芽密度、叶片着生状态与叶缘的信息。其中,前5个主成分累积方差贡献率(53.32%)在50%以上,反映了茶树种质资源的大部分信息[54],说明叶宽、叶长/叶宽、百芽重、一芽三叶长、节间长、梗粗/节间长、叶长等性状是引起茶树种质表型差异的主要因素。
2.4.2 综合评价 按照PCj(j=1~10)和相应特征向量及茶树种质资源各性状标准化值,计算出72份茶树种质10个主成分值Z(i,j)(i=1~72),然后依据10个主成分值及其对应特征根值建立线性方程:Zi=0.219×Z(i,1)+0.150×Z(i,2)+0.143×Z(i,3)+ 0.105×Z(i,4)+0.084×Z(i,5)+0.075×Z(i,6)+0.064×Z(i,7)+ 0.057×Z(i,8)+0.054×Z(i,9)+0.049×Z(i,10),计算各种质的综合得分(表1)。结果显示,WL06、WL44、WL41、WL72和WL26的综合得分均大于1,排名前5,WL64和WL59均小于–1,排名后2。排名前5与排名后2的茶树种质在百芽重、叶宽、一芽三叶长、梗粗、芽叶色泽、叶长/叶宽值等重要性状上存在明显差异。研究表明,芽叶色泽紫红、紫绿色或黄绿色[16]及叶片宽、叶长/叶宽值小、梗粗的鲜叶有利于乌龙茶品质的形成[17],百芽重、发芽密度可用于间接判断茶树的产量[18]。据此推测,WL06、WL44、WL41、WL72和WL26在乌龙茶的品质和产量方面表现优良。
3 讨论
种质资源的遗传多样性是茶树新品种成功选育及应用方面突破的关键,对其进行分析和保护,既拓宽了茶树品种改良的遗传基础,保证长期稳定的改良效果,也可以根据产业的需求变化培育新品种[55]。本研究结果显示,27项表型性状的遗传多样性指数(H')在0.51~2.15之间,H'平均值为1.30,明显高于福建武夷山(H'=1.09)[22]、陕西(H'=0.97,1.11)[35-36]、云南(H'=1.04,0.95)[39,40]、贵州(H'=1.11)[41]、河南(H'=0.97)[43]等地方茶树种质资源,说明来源于不同产地的茶树种质资源能够拓宽茶树品种改良的遗传基础,表现出丰富的遗传多样性,这与黃福平等[29]、段云裳等[30]、Lin等[31]在分子水平上的研究结果基本一致。数量性状的H'平均值(1.82)远大于质量性状(0.94),说明不同来源地茶树种质资源数量性状的遗传基础广、变异类型丰富、分布均匀,具有较大的遗传改良潜力。数量性状的变异系数均大于10%[56],在茶树种质个体间均存在较大的差异,说明不同茶树种质间在数量性状上的变异程度不一致,可拓宽茶树新品种选育的空间和方向。上述分析表明,参试的茶树种质资源表型多样性丰富,可为新品种的选育和杂交亲本的选择提供丰富的种质材料。
相关性分析可以明确表型性状之间的影响关系,可为茶园的田间管理和品种形态性状的遗传改良提供参考。本研究结果显示,不同来源地茶树种质资源多个数量性状间存在着复杂的相关性,其中叶长与叶宽,叶长、叶宽与侧脉对数、一芽三叶长、花冠直径均呈极显著正相关,叶宽与节间长、梗粗显著正相关,叶宽与发芽密度、叶长/叶宽显著或极显著负相关,这与陕西[36]、云南[40]、江苏[42]等地茶树种质资源研究结果基本一致。此外,百芽重与叶长、叶宽等极显著正相关,与叶长/叶宽等显著或极显著负相关;发芽密度与梗粗/节间长显著正相关,与百芽重等显著或极显著负相关;一芽三叶长与节间长、花冠直径等极显著正相关,与梗粗/节间长极显著负相关;梗粗与节间长等显著或极显著正相关,与叶长/叶宽显著负相关;花冠直径与百芽重等极显著正相关,与发芽密度等显著负相关。因此,依据性状间的相关性,可以根据育种目标选择亲本材料进行多性状的遗传改良,还可以根据生产目的采取科学的栽培管理措施进行性状的直接或间接调控,提高生产效率及茶树鲜叶的产量、质量。
基于27项表型性状的聚类分析结果显示,72份茶树种质在遗传距离为16时被划为4个类群,其中来自福建和台湾的茶树种质资源在表型上的亲缘关系较近,二者均与来自广东的种质资源间存在较远的亲缘关系,该结果与黄福平等[29]、段云裳等[30]、Lin等[31]的研究结果相吻合。不同类群间在芽叶色泽、百芽重、一芽三叶长、节间长、梗粗、梗粗/节间长、叶长、叶宽、叶长/叶宽等性状上差异显著甚至极显著,说明72份茶树种质资源的遗传聚类与这些性状关系密切。研究认为,树型、树姿及花冠直径的变化是茶树演化的主要形态变化[18],据此推测,第Ⅳ类群为半野生型:树型小乔木型、树姿多直立、花冠直径大;第Ⅰ、Ⅱ类群为混合型:树型为小乔木或灌木型,树姿直立、半开张或开张,花冠直径居中;第Ⅲ类群为栽培型:树型为灌木型、树姿多半开张或开张、花冠直径小。基于表型性状明确了各类群间的形态差异、形态特点与进化类型,可为不同来源地茶树种质资源的遗传进化研究提供参考。
主成分分析是把原来多个具有一定相关性的变量划为少数几个互不相关的综合指标,是一种降维处理技术[57],该方法可简化选择程序、获得解释方差的重要性状[58],在茶树[22-24,47]种质资源的鉴定评价中应用效果良好。本研究结果显示,前10个主成分代表了27项表型性状绝大多数信息,可以用来对不同来源地茶树种质资源进行综合评价[59]。依据综合得分大小,筛选出WL06、WL44、WL41、WL72、WL26等5份茶树种质在茶树产量、乌龙茶品质等方面表现良好。其中,WL06和WL41的鉴定结果与其在福建福安的表现比较一致[2],WL44的鉴定结果与曹士先等[60]的研究结果比较一致。因此,这些表型优异的茶树种质,今后既可作为茶叶新产品开发、新品种选育或杂交亲本直接加以利用,又可作为茶树遗传改良中优良基因挖掘利用的研究材料。
综上认为,来自福建、广东、台湾的茶树种质资源组成的群体,其表型多样性丰富,个体间数量性状差异较大,并存在复杂的相关性;通过聚类分析揭示了不同来源地茶树种质资源间的亲缘关系,明确了4个群体的表型差异、形态特点与进化类型;基于主成分分析和综合得分,筛选出用于评价茶树亲本的重要形态指标和5份综合性状优良的茶树种质。在同一生境和田间管理条件下,本试验取得的研究结果具有一定的真实性、可靠性,可为茶树新品种选育、杂交亲本选择及种质资源的开发利用提供参考。但基于表型性状对茶树种质资源进行鉴定评价与遗传多样性分析,仍具有一定的局限性,因此在此基础上,今后可借助其他技术手段对其进行深入研究,以期获得更加全面直接的研究结果。
参考文献
[1] 陈荣冰, 王飞权. 乌龙茶种质资源创新与应用研究[M]. 北京: 中国原子能出版社, 2019.
[2] 陈常颂, 余文权. 福建省茶树品种志[M]. 北京: 中国农业科学技术出版社, 2016: 32-66, 84-104.
[3] 罗盛财, 陈德华, 黄贤格, 等. 武夷名丛单丛茶树种质资源收集、整理鉴定与保护利用研究[J]. 中国茶叶, 2017, 39(12): 18-20.
[4] 陈伟忠, 陈若荣, 陈思藩, 等. 凤凰单丛茶树种质资源保存及利用现状和展望[J]. 亚热带农业研究, 2016, 12(3): 209-215.
[5] 陈荣冰, 姚信恩. 台湾乌龙茶品种[J]. 中国茶叶, 2002(4): 10-11.
[6] 游小妹, 陈常颂, 林郑和, 等. 金萱杂交创新种质的初步筛选鉴定[J]. 茶叶科学技术, 2011(4): 19-23.
[7] 林郑和, 游小妹, 钟秋生, 等. 高香型茶树新品系初步筛选[J]. 福建农业学报, 2014, 29(3): 227-232.
[8] 陈志辉, 游小妹, 林郑和, 等. 茶树新品种 ‘瑞茗 全国区域试验报告[J]. 茶叶学报, 2020, 61(3): 101-108.
[9] 周李华. 广东茶树种质资源遗传多样性AFLP分析[D]. 长沙: 湖南农业大学, 2006.
[10] 丁帅涛, 程晓梅, 张 亚, 等. 基于表型性状和生化成分的陕西茶树种质资源遗传多样性研究[J]. 西北农业学报, 2019, 28(4): 607-619.
[11] Chen X J, Wang P J, Zheng Y C, et al. Comparison of metabolome and transcriptome of flavonoid biosynthesis pathway in a Purple-leaf tea germplasm Jinmingzao and a Green-leaf tea germplasm Huangdan reveals their relationship with genetic mechanisms of color formation[J]. International Journal of Molecular Sciences, 2020, 21(11): 4167.
[12] Cheng S H, Fu X M, Liao Y Y, et al. Differential accumulation of specialized metabolite L-theanine in green and albino-induced yellow tea (Camellia sinensis) leaves[J]. Food Chemistry, 2019, 276: 93-100.
[13] Guo Y, Chang X, Zhu C, et al. De novo transcriptome combined with spectrophotometry and gas chromatography -mass spectrometer (GC-MS) reveals differentially expressed genes during accumulation of secondary metabolites in purple-leaf tea (Camellia sinensis cv Hongyafoshou)[J]. The Journal of Horticultural Science and Biotechnology, 2019, 94(3): 349-367.
[14] Li C F, Ma J Q, Huang D J, et al.Comprehensive dissection of metabolic changes in albino and green tea cultivars[J]. Journa of Agricultural and Food Chemistry, 2018, 66(8): 2040-2048.
[15] Wei K, Wang L Y, Zhou J, et al. Comparison of catechins and purine alkaloids in albino and normal green tea cultivars (Camellia sinensis L.) by HPLC[J]. Food Chemistry, 2012, 130(3): 720-724.
[16] 郭吉春, 葉乃兴, 何孝延, 等. 乌龙茶品种资源研究进展[C]//中国茶叶学会、台湾大学食品科技研究所、福建茶叶进出口有限责任公司. 海峡两岸茶叶科技学术研讨会论文集, 2000.
[17] 宛晓春. 茶叶生物化学[M]. 3版. 北京: 中国农业出版社, 2003: 248-249.
[18] 江昌俊. 茶树育种学[M]. 2版. 北京: 中国农业出版社, 2011: 12-13, 82-84.
[19] 王飞权, 冯 花, 王 芳, 等. 42份武夷名丛茶树资源生化成分多样性分析[J]. 植物遗传资源学报, 2015, 16(3): 670-676.
[20] 张见明. 26份武夷名丛茶树种质资源秋茶生化成分多样性分析[J]. 武夷学院学报, 2016, 35(9): 1-7.
[21] Chen S, Li M H, Zheng G Y, et al. Metabolite profiling of 14 Wuyi rock tea cultivars using UPLC-QTOF MS and UPLC-QqQ MS combined with chemometrics[J]. Molecules, 2018, 23(2): 104-122.
[22] 王飞权, 冯 花, 罗盛财, 等. 武夷名丛茶树种质资源农艺性状多样性分析[J]. 中国农业科技导报, 2019, 21(6): 43-54.
[23] 王飞权, 李纪艳, 冯 花, 等. 武夷名丛茶树种质资源叶片解剖结构分析[J]. 热带作物学报, 2019, 40(12): 2375-2389.
[24] 石玉涛, 郑淑琳, 王飞权, 等. 武夷名丛茶树种质资源矿质元素含量特征分析[J]. 中国农业科技导报, 2020, 22(7): 37-50.
[25] 叶江华, 罗盛财, 王海斌, 等. 武夷山名丛单丛茶树种质资源的遗传多样性与亲缘关系分析[J]. 福建茶叶, 2017, 39(5): 11-13.
[26] 陈志丹, 林志坤, 孙威江, 等. 125份武夷山茶树种质资源的遗传多样性分析及DNA指纹图谱的建立[J]. 分子植物育种, 2016, 14(12): 3601-3608.
[27] 陈志丹, 李振刚, 孙威江. 闽南乌龙茶茶树种质资源RAPD指纹图谱构建及遗传多样性分析[J]. 分子植物育种, 2012, 10(6): 731-739.
[28] 晏嫦妤. 凤凰单丛古茶树资源遗传多样性AFLP分析及保护研究[D]. 长沙: 湖南农业大学, 2007.
[29] 黄福平, 梁月荣, 陆建良, 等. 乌龙茶种质资源种群遗传多样性AFLP评价[J]. 茶叶科学, 2004(3): 183-189.
[30] 段云裳, 成 浩, 姜燕华, 等. 乌龙茶品种(系)遗传多样性与亲缘关系的SSR分析[J]. 茶叶科学, 2010, 30(2): 141-148.
[31] Lin Y, Yu W T, Zhou L, et al. Genetic diversity of oolong tea (Camellia sinensis) germplasms based on the nanofluidic array of single-nucleotide polymorphism (SNP) markers[J]. Tree Genetics & Genomes, 2020, 16(1): 3-17.
[32] 李洪果, 陈达镇, 许靖诗, 等. 濒危植物格木天然种群的表型多样性及变异[J]. 林业科学, 2019, 55(4): 69 -83.
[33] 張 叶, 叶蓓蕾, 邬 静, 等. 77份文心兰种质资源表型性状遗传多样性分析[J]. 热带作物学报2021, 42(8): 2183-2190.
[34] 王小萍, 张 厅, 黄 梅, 等. 四川地区茶树种质资源叶表型性状多样性分析[J]. 中国农学通报, 2018, 34(4): 89-94.
[35] 班秋艳, 纪晓明, 余有本, 等. 陕西茶树种质资源表型性状的遗传多样性研究[J]. 安徽农业大学学报, 2018, 45(4): 575-579.
[36] 丁帅涛, 程晓梅, 张 亚, 等. 陕西古茶树种质资源表型性状遗传多样性研究[J]. 广东农业科学, 2018, 45(2): 52-60, 2.
[37] 郑旭霞, 黄海涛, 沈林章, 等. 紫笋群体种茶树资源表型性状特性和遗传多样性分析[J]. 茶叶, 2020, 46(4): 223-228.
[38] 刘福桥, 李 强, 戎玉廷, 等. 云南双江县古茶树种质资源的表型多样性[J]. 中国茶叶, 2017, 39(4): 22-25.
[39] 蒋会兵, 宋维希, 矣 兵, 等. 云南茶树种质资源的表型遗传多样性[J]. 作物学报, 2013, 39(11): 2000-2008.
[40] 陶 萍, 宋 燚, 张晴晖, 等. 云南普洱茶种古茶树资源表型性状数据分析研究[J]. 西部林业科学, 2020, 49(6): 144-151, 157.
[41] 牛素贞, 宋勤飞, 安红卫, 等. 贵州古茶树种质资源基于形态特征的多样性研究[J]. 浙江农业学报, 2019, 31(10): 1689-1699.
[42] 俞文生, 倪佳成, 陈 慧, 等. 茶树种质资源表型遗传多样性分析[J]. 安徽农业科学, 2017, 45(25): 40-42, 50.
[43] 潘宇婷, 袁正仿, 袁红雨, 等. 河南省地方茶树种质资源表型性状遗传多样性研究[J]. 信阳师范学院学报(自然科学版), 2018, 31(4): 578-585.
[44] 陈 杏, 农玉琴, 廖春文, 等. 基于表型性状的广西茶树种质资源遗传多样性分析[J]. 中国热带农业, 2019(4): 49-52.
[45] 丁 洲, 李烨昕, 袁 艺, 等. 安徽茶区优良群体种的表型性状和遗传多样性分析[J]. 茶叶科学, 2018, 38(2): 155-161.
[46] 王治会, 彭 华, 江新凤, 等. 江西茶树种质资源芽叶表型多样性分析[J]. 江苏农业科学, 2020, 48(1): 134-138, 142.
[47] 郑淑琳, 石玉涛, 王飞权, 等. 乌龙茶种质资源矿质元素含量特征分析与评价[J]. 福建农业学报, 2020, 35(2): 150-160.
[48] 陈 亮, 虞富莲, 杨亚军, 等. 农作物种质资源鉴定技术规程 茶树: NY/T1312-2007[S]. 北京: 中国农业出版社, 2007.
[49] 陈 亮, 虞富莲, 杨亚军, 等. 茶树种质资源与遗传改良[M]. 北京: 中国农业科学技术出版社, 2006: 63-97.
[50] 余文权, 陈常颂. 福建茶树良种选育与应用[M]. 北京: 中国农业科学技术出版社, 2015: 1-25.
[51] 王新超, 陈 亮, 杨亚军. 广西茶树资源生化成分多样性分析[J]. 植物遗传资源学报, 2010, 11(3): 309-314, 319.
[52] 石 林. 聚类分析在茶树品种分类中的应用[J]. 中国茶叶, 1988(3): 15-17.
[53] Diederichsen A, Raney J P, Duguid S D. Variation of mucilage in flax seed and its relationship with other seed characters[J]. Crop Science, 2006, 46(1): 365-371.
[54] 匡立學, 聂继云, 李志霞, 等. 不同苹果品种果实矿质元素含量的因子分析和聚类分析[J]. 中国农业科学, 2017, 50(14): 2807-2815.
[55] 王明麻. 林木遗传育种学[M]. 北京: 中国林业出版社, 2001.
[56] 孙 铭, 符开欣, 范 彦, 等. 15份多花黑麦草优良引进种质的表型变异分析[J]. 植物遗传资源学报, 2016, 17(4): 655-662.
[57] 季彪俊. 田间试验的SPSS统计分析[M]. 北京: 中国农业出版社, 2014: 201-214.
[58] 王建丽, 刘杰淋, 朱瑞芬, 等. 28份籽粒苋种质资源的主要农艺性状遗传多样性分析[J]. 草地学报, 2020, 28(4): 1050-1059.
[59] 李清华. 34份菜用大豆品种主要农艺性状的主成分分析及遗传距离测定[J]. 福建农业学报, 2018, 33(2): 136-143.
[60] 曹士先, 冯卫虎, 林燕清, 等. 茶树品系“金凤凰”早期鉴定及试制性研究初探[J]. 福建农业, 2017(8): 40-42.
责任编辑:沈德发
