不同水深对挺水植物形态和生理指标的影响
2021-11-22黄晓龙谢洪民魏伟伟李宽意
黄晓龙, 谢洪民, 魏伟伟, 李宽意,3,4
1.中国科学院南京地理与湖泊研究所, 湖泊与环境国家重点实验室, 江苏 南京 210008
2.中国环境科学研究院, 湖泊水污染治理与生态修复技术国家工程实验室, 北京 100012
3.中国科学院大学中丹学院, 北京 100049
4.重庆三峡学院环境与化学工程学院, 重庆 404000
影响水生植物生长、繁殖和分布的因素包括生物因素和非生物因素,生物因素主要包括藻类、浮游生物、鱼类、附着生物和种内种间竞争等;而起到决定性作用的则是非生物因素,包括水深、pH、光照、营养元素、透明度和基质类型等[1-2]. 其中,水深是对水生植物生长繁殖起关键作用的生态因子,但对4种水生植物生活型(漂浮植物、沉水植物、浮叶植物和挺水植物)的影响并不一致. 漂浮植物生长在水体表面,较少受水深波动的影响. 沉水植物和浮叶植物可通过补偿生长调节株高、节间距、地上地下生物量分配和形成异型叶等方式调节对水深变化的适应,因此水深对它们的影响较小[3-6]. 但挺水植物对水深的适应性较低,水深是其生长最重要的限制因子,较低水深会限制挺水植物生长,较高水深会淹没植株,造成植物根部缺氧,导致挺水植物死亡[7-10].
挺水植物是湖滨沿岸带湿地系统重要组成部分,其部分叶片和茎杆挺出水面,根系或地下茎生于底质中,有发达的通气组织,主要分布在水深较浅的沿岸带,兼具有陆生和水生植物的特点,被视作植物从陆生到水生的过渡类型[11-14]. 挺水植物可形成具层次的湿地景观,在人工湿地构建中起到重要作用,其初级生产力在4种水生植物生活型中往往也是最高的[15-16]. 挺水植物可直接从基质中吸收营养元素,对生活污水内污染物的吸附能力也较强,可以有效削减流入湖泊中的外源营养元素负荷,收割其地上部分能够去除吸收的氮磷等营养元素[17-18];同时,其根系固着于湖泊沿岸带底质中,可减弱风浪对沿岸带的冲击,间接减少营养元素的释放.
太湖流域河网密布,湖泊众多,是中国乃至全球重要的湿地分布区,具有蓄纳洪水、调节地表径流的重要水文功能. 然而,目前太湖流域面临多重自然和人为因素的威胁[19-20],其湖泊一般平均深度低于 3 m,是一类较为脆弱的淡水生态系统[21]. 作为初级生产者,水生植物在淡水生态系统中为其他营养级的生物提供栖息地和繁殖场所,其果实、种子、根状茎和块茎也是鸟类喜爱的食物[4,8,12]. 目前,受灌溉、水污染、水利工程、岸带固化、房地产开发建设等诸多直接或间接人类活动干扰,造成了包括水生植物在内的淡水生态系统多样性下降、结构简化、生态服务功能下降的问题[22-24]. 经过中国环境科学研究院研究团队前期对太湖流域水生植物调查,通过“四重筛选法”筛选得到适宜太湖流域地区沿岸带水生植物的初筛种、适应河口沿岸带水力冲击和水深变化的次筛种、对沿岸带河口水质具有明显改善效果的三筛种,以及真实沿岸带生境下对水力冲击适应性高、对透明度改善效果明显、对氮磷去除效果好且衰亡分解速率低的四筛种(终选种)水生植物,即菰(Zizanialatifolia)、慈姑(Sagittariatrifolia)、再力花(Thaliadealbata)和苦草(Vallisnerianatans)[25-27]. 其中,苦草为沉水植物,不在该研究的范围内. 该研究拟根据太湖流域沿岸带实际情况设置不同梯度水深高度〔(高水深(40 cm)、中水深(20 cm)和低水深(10 cm)〕,培植前3种挺水植物(菰、慈姑和再力花),测定叶片和根系形态指标、相对生长速率指标和生理指标,探究不同水深对挺水植物形态和生理指标的影响. 该研究有助于在太湖流域生态修复工程中针对不同水深变化配置挺水植物组合.
1 材料与方法
1.1 研究区概况
该研究于2019年9月17日—10月29日在太湖湖泊生态系统国家野外科学观测研究站(简称“太湖站”)进行. 太湖站(31.419 8°N、120.215 9°E)隶属于中国生态系统观测研究网络(CNERN),是唯一一个加入全球湖泊生态观测网络(GLEON)的中国成员单位. 太湖站位于江苏省无锡市滨湖区太湖沿岸带上,属亚热带季风性气候,光照充足,降水充沛,雨热同期. 太湖为我国长江下游地区最大的淡水湖,面积约 2 338 km2,是我国第三大淡水湖,湖底平均高程仅1.1 m(吴淞高程),平均水深1.9 m,最大水深2.6 m,目前作为一个亚热带季风区典型浅型淡水湖泊被广泛研究[28-30]. 太湖流域是中国经济最发达的地区之一,虽然流域面积只占我国领土面积的1/100,人口只占我国总人口的1/22,但2019年贡献了我国1/10的国内生产总值(GDP)[31].
1.2 植物材料
试验开始前,在太湖沿岸带野外采集菰、再力花和慈姑3种挺水植物幼苗各30株,带回太湖站内培养. 菰,又称茭草,为禾本科菰属多年生植物,具发达地下茎,茎高而直立,成株高1~2 m,可在10~25 ℃的合适温度下生长. 菰为长江中下游地区湖泊沿岸带优势植物[32],对日照长短要求不严格,对水的需求高,对水深变化敏感,在水深较高的湖区地下茎无法萌发消亡,在水深较低地区则过度繁殖,成为优势群落,导致湖泊沼泽化.
慈姑,又称茨菇、野慈姑,为泽泻科慈姑属多年生植物,具粗壮根状茎,叶片剑形,适应能力强,为中国广布种[33-34].
再力花,又称水竹芋,竹芋科再力花属,具较高观赏价值,根系发达,净化水体能力强,作为引入植物广泛用于富营养化水体生态修复[35-36]. 虽然现在尚不认为其具备入侵危害,但这种植物在我国尚未经过入侵风险评价[37].
1.3 试验设计
该试验开始于2019年9月17日,设置双因素试验,因素为植物和不同水深高度. 试验在太湖站室外聚乙烯试验桶(直径0.46 m,高0.90 m,容积约150 L)中进行. 试验开始前采集太湖湖泥〔w(TN)为1.73 mg/g,w(TP)为0.4 mg/g〕,混匀后放置15 cm高底泥于试验桶中. 试验开始前挑选大小一致的植株,测定植物初始根茎干质量. 试验用水为过40目(孔径0.450 mm)滤网的太湖湖水,水体指标(见表1)均使用美国YSI公司Pro1030多参数水质分析仪测定. 太湖沿岸带挺水植物一般分布于水深约0.8 m以内的范围[28],但现场调查发现在0.8 m水深下只有芦苇能够生存. 根据在太湖沿岸带调查的实际情况,设置水深高度为低水深(10 cm)、中水深(20 cm)、高水深(40 cm)(均从湖泥表面算起). 每个处理重复4次,共使用36(3×3×4)个试验桶. 试验期间控制水深在设置高度,当天气预报有降水时使用桶盖覆盖试验桶,蒸发强烈时用上述过滤后湖水补充至相应水深高度.

表1 试验期间水体指标
试验开始时植物形态指标如表2所示. 由表2可见,3种挺水植物的干质量无显著性差异,但叶面积和根系长度的差异性均显著,其大小均表现为慈姑>再力花>菰.

表2 试验开始时植物形态指标
该试验于2019年10月29日结束,历时42 d,此时不同水深植物间出现明显差异. 收获时将试验桶中水排出,倾斜试验桶小心铲出基质,用水仔细清洗去除根系上附着的基质. 剪开地上部分(茎干和叶片)和地下部分(根系和地下茎). 植物叶片和根系先使用佳能LiDE 220扫描仪(美国)扫描,采用ImageJ软件分析叶片和根系数据后,再放于65 ℃烘箱中烘干至恒质量,最后测定植物地上和地下部分干质量、叶面积、根系长度以及相对生长速率(relative growth rate,RGR). 相对生长速率计算公式:
RGR=ln(r2/r1)/t
(1)
式中:r1和r2分别为试验前、后植物地上部分和地下部分干质量之和,g;t为试验时间,该研究取值42 d.
试验收获时取植物最高处倒数第二片新鲜叶片测定植物超氧化物歧化酶(superoxide dismutase,SOD)和丙二醛(malonic dialdehyde,MDA)含量. 取植物最高处倒数第3片叶片65 ℃烘干24 h后进行总可溶性糖含量测定. 生理指标的测定均使用UV-Vis 2450紫外-可见光分光光度计(日本岛津公司)测定,具体参照《植物生理学实验指导》[38]. 总可溶性糖含量采用蒽酮-硫酸法,在625 nm波长下并使用葡萄糖溶液做标准曲线测定,单位为μg/g;SOD含量采用氮蓝四唑光还原法,在560 nm波长下测定,以能抑制50%反应酶为一个SOD反应酶单位(U),单位为U/g;MDA含量采用硫代巴比妥酸法测定,在532和450 nm波长下测定,单位为nmol/g.
1.4 数据分析
在统计分析之前分别通过Shapiro-Wilk检验和Levene检验确定植物叶片和根系形态指标、相对生长速率指标和生理指标是否满足正态性和方差齐次性假设. 如果数据不能满足正态性或方差齐次性假设,那么这组指标和对应的指标均进行lg(x+1)转换以达到数据分析要求. 植物和水深作为两个因子进行对植物形态和生理指标的双因素方差分析(two-way ANOVA),之后使用Duncan检验进行指标间两两比较(显著性阈值设置为P=0.05). 采用SPSS 26软件进行数据分析;使用SigmaPlot 14 软件进行绘图.
2 结果与分析
在不同水深高度对植物性状指标的影响试验中,植物和水深作为双因素方差分析的2个主因素,分析结果如表3所示. 由表3可知,除MDA指标外,在P<0.05水平上不同植物间的差异显著. 不同水深对植物性状指标均有显著影响. 植物和水深在MDA上的交互作用显著,其他性状指标则不存在显著交互作用.

表3 植物和水深作为两个因子对植物性状指标的双因素方差分析
2.1 挺水植物地上部分对水深的响应
图1为不同水深高度对3种挺水植物地上部分干质量的影响. 由图1可知:在低水深条件下慈姑地上部分干质量最高,显著高于中水深和低水深条件,并且在低水深、中水深、高水深条件下均高于再力花和菰;在中水深条件下,再力花地上部分干质量相对低水深和高水深条件均较低;菰地上部分干质量则随水深由高向低逐渐降低.

注: 相同小写字母表示不同处理间无显著性差异(P>0.05),不同小写字母表示不同处理间存在显著性差异(P<0.05).下同.
图2为不同水深高度对3种挺水植物叶面积的影响. 由图2可知:在低水深条件下慈姑叶面积最高,显著高于中水深和低水深条件,并且在低水深、中水深、高水深条件下均高于再力花和菰;在中水深条件下再力花叶面积最高,且高水深和低水深条件差异不大;在高水深条件下菰叶面积最高,且低水深和中水深条件差异较小.

图2 不同水深高度对3种挺水植物叶面积的影响
2.2 挺水植物地下部分对水深的响应
图3为不同水深高度对3种挺水植物地下部分干质量的影响. 由图3可知:在低水深条件下慈姑地下部分干质量最高,显著高于中水深和低水深条件,并且在低水深、中水深、高水深条件下均高于再力花和菰;在中水深处条件下再力花地下部分干质量最高,高水深条件次之,低水深条件最低;菰地下部分干质量在高水深和低水深条件下的差异不大,但均高于中水深条件.

图3 不同水深高度对3种挺水植物地下部分干质量的影响
图4为不同水深高度对3种挺水植物根系长度的影响. 由图4可知:在低水深条件下慈姑根系长度最高,显著高于中水深和低水深条件,并且在低水深、中水深、高水深条件下均高于再力花和菰;在中水深条件下再力花根系长度最高,且高水深和低水深条件差异不大;在高水深条件下菰根系长度最高,低水深条件次之,中水深条件最低.
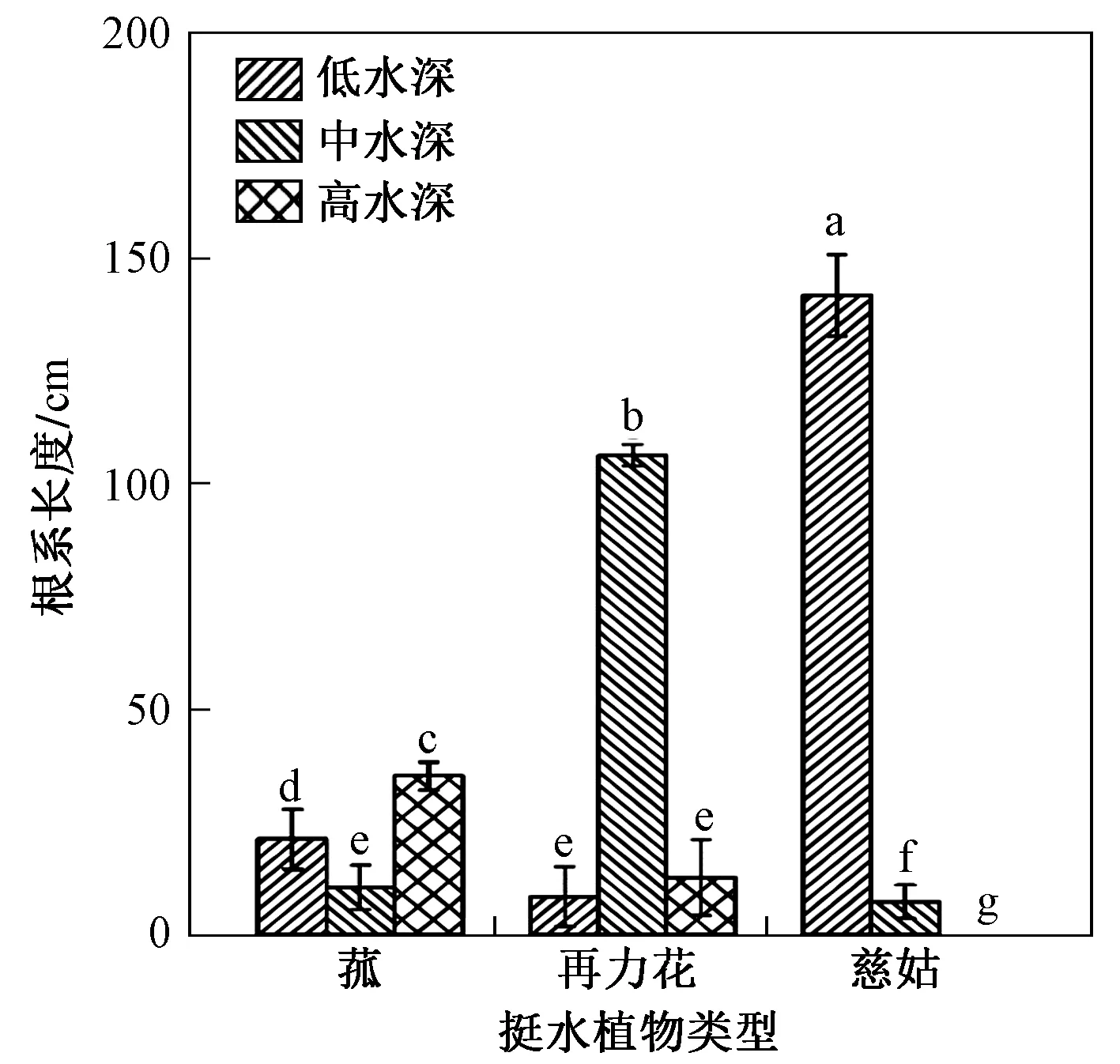
图4 不同水深高度对3种挺水植物根系长度的影响
2.3 挺水植物生长指标对水深的响应
图5为不同水深高度对3种挺水植物相对生长速率的影响. 由图5可知:在低水深条件下慈姑表现出较高的相对生长速率,且随水深增加呈逐渐递减趋势;在中水深条件下再力花的相对生长速率最高,显著高于低水深和高水深条件,且低水深和高水深条件差异不显著;慈姑在低水深条件下表现出较高的相对生长速率,显著高于中水深和高水深条件,且中水深和高水深条件差异不大.
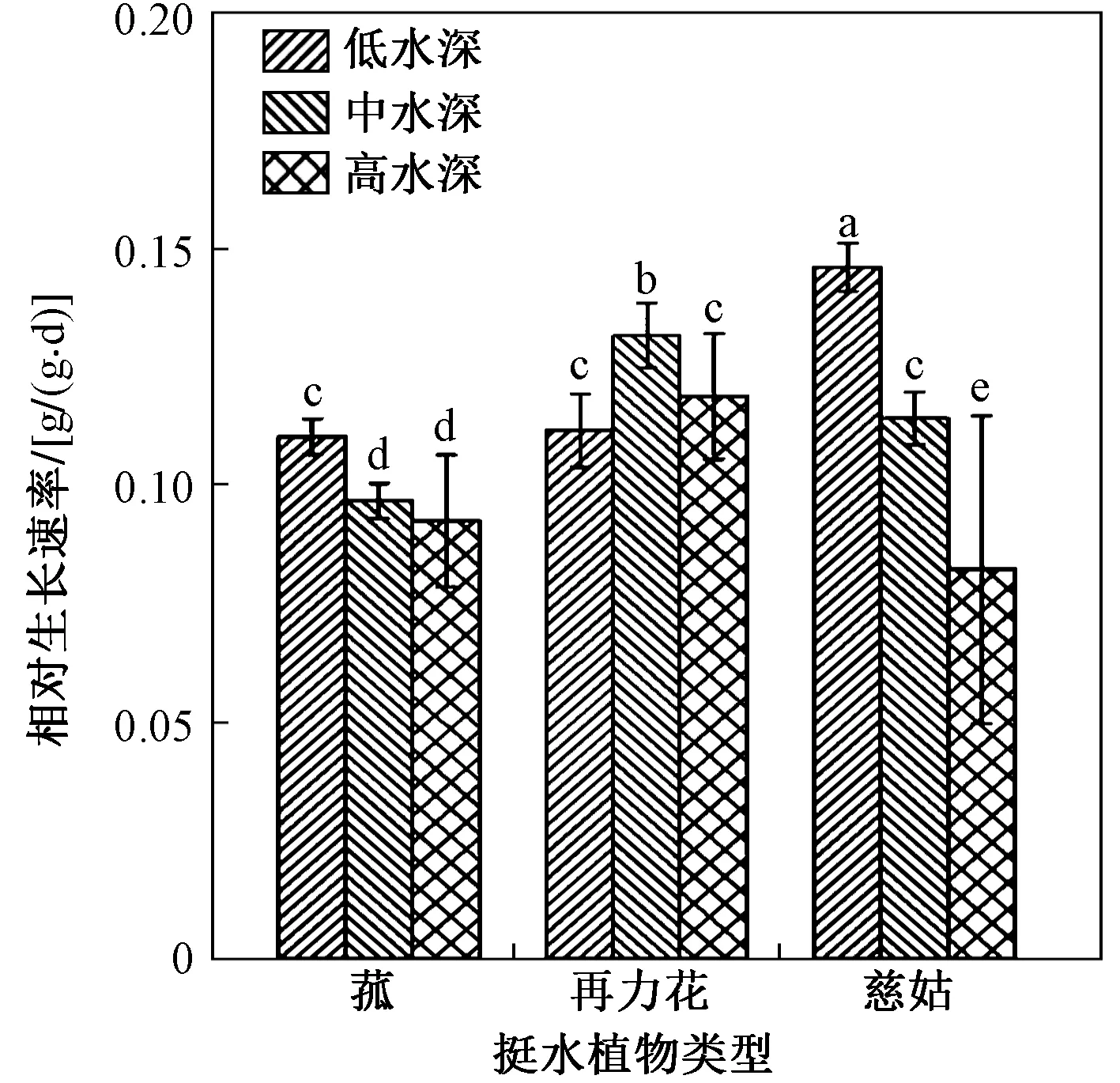
图5 不同水深高度对3种挺水植物相对生长速率(RGR)的影响
2.4 挺水植物生理指标对水深的响应
图6为不同水深高度对3种挺水植物生理指标总可溶性糖含量的影响. 由图6可知,随水深升高,3种挺水植物的总可溶性糖含量均呈上升趋势. 在低水深条件下,3种挺水植物的总可溶性糖含量均较低,其中菰的总可溶性糖含量相对较高;在中水深条件下,菰的可溶性糖含量较高;在高水深条件下,3种挺水植物的总可溶性糖含量大小表现为慈姑>再力花>菰.

图6 不同水深高度对3种挺水植物生理指标总可溶性糖含量的影响
图7为不同水深高度对3种挺水植物生理指标超氧化物歧化酶(SOD)含量的影响. 由图7可知,随水深升高,3种挺水植物的SOD含量均呈上升趋势. 在低水深条件下,3种挺水植物的SOD含量均较低,其中慈姑SOD含量最低;在中水深和高水深条件下,SOD含量大小均表现为慈姑>再力花>菰.

图7 不同水深高度对3种挺水植物生理指标超氧化物歧化酶(SOD)含量的影响
图8为不同水深高度对3种挺水植物生理指标丙二醛(MDA)含量的影响. 由图8可知,随水深升高,3种挺水植物的MDA含量均呈上升趋势,且再力花在中水深和高水深条件下的差异不显著. 在低水深条件下,3种挺水植物均表现出较低的MDA,其中慈姑MDA含量最低;在中水深和高水深条件下,MDA含量大小均表现为慈姑>再力花>菰.

图8 不同水深高度对3种挺水植物生理指标丙二醛(MDA)含量的影响
3 讨论
慈姑虽然在试验收获时地上部分干质量较高,但主要残留器官为挺立的植物茎干,由于叶片均已脱落,叶面积接近于0(见图2),生理指标也呈现补偿性增长的趋势(见图6~8),显示高水深对慈姑具有较强的抑制作用. 再力花也呈现出相近的趋势,但对水深的耐受性高于慈姑,虽然高水深抑制了再力花的生长,在中水深条件下仍呈现较明显的增长趋势. 再力花为外来种,目前尚未经过外来种入侵风险评估,在园林绿化中使用比较广泛,这是由于这种植物虽然是外来种,但已经在太湖流域生长多年,适应性强,效果好,且可以兼顾一定的景观功能;实践证明,再力花不具备强大和快速的繁殖能力,入侵风险较小,但在太湖沿岸带水生植物恢复中仍应慎重使用. 菰对高水深的适应性较强,虽然高水深也抑制了地上部分生长,但叶面积却呈现显著增加趋势(见图1、2),显示出补偿生长的趋势[39]. 通过该试验结果可见,菰对高水深较为适应,这与王慧丽等[32]研究结果相似.
该研究中3种挺水植物生理指标对高水深均表现出较为一致的响应性. 在高水深这一胁迫条件下,植物倾向于产生更多的总可溶性糖、SOD和MDA,显示出植物对高水深这一胁迫的生理适应性(见图6~8). 总可溶性糖为植物重要的生长指标,它是植物生长和繁殖最主要的能量来源,同时也是植物合成其他化合物的原料. 在该研究中,受高水深胁迫的情况下,挺水植物倾向于产生更多的总可溶性糖,用以提高对不良环境的抵抗性,这与马佳等[18]研究结果一致. 植物叶绿体中的SOD一般为Fe-SOD,它通过催化O2-自由基生成O2和H2O2,较高的SOD表明植物具有较高的抗胁迫能力. MDA为植物自由基作用于脂类物质发生过氧化反应的产物,植物受到胁迫时对丙二醛的积累增加,同样是表征植物抗逆性的一个指标. 虽然菰对高水深的叶片和根系形态指标、相对生长速率指标表现均较好,但仍表现出较高的生理指标,显示虽然高水深没有抑制菰的生长,但仍然对其生长进行了胁迫.
湖泊沿岸带处于陆地和水体的接触区,多风浪,水土流失严重,生态系统功能脆弱,受人类干扰较重[40-42]. 太湖沿岸带由于受到人为强烈干扰,水生植被,尤其是挺水植物植被严重破坏. 虽然这一地区水生植物收割已经被禁止,但是水生植物覆盖面积下降仍是事实[19]. 这一地区也面临水生植物优势种由本地种转变为外来种、本地种多样性下降的趋势[43-44]. 1960年挺水植物占太湖水生植物总生物量的80%,然而到2014年,挺水植物仅占太湖水生植物总生物量的10%[45]. 由于在生长期内连续面临干旱和淹水胁迫,在湖泊沿岸带进行生态修复比陆地或水体内更困难. 推进湖泊沿岸带生态修复,筛选适合的挺水植物是目前的技术难题. 常规的生态修复方案往往只选用一种植物,好处是易于培育,种植方便,不足之处在于多样性较差、无法抵抗水深变化、不具层次性的景观[46].
挺水植物茎叶挺出水面,水深升高会直接限制挺水植物的光合作用,此外水深还会影响植物根系的氧化还原情况. 植物处于淹水胁迫下,导致根系长时间处于缺氧或厌氧环境,在淹水条件下降低底质的氧化还原电位,导致有毒有害物质积累,限制了植物的生长[1,3]. 不同挺水植物对水深高度的响应不同,每种植物都有一个最适宜的水深[10]. 挺水植物也可以通过形态上延长茎间距离、增加植株高度来缓解淹水胁迫[46-47]. 在该研究中针对水深胁迫,挺水植物通过调节形态和生理指标来适应变化,抵抗水深胁迫带来的不利影响. 可以根据该研究结果在不同水深高度范围内栽种挺水植物,这有助于该地区水生植物的恢复.
4 结论
a) 不同水深对挺水植物性状指标均有显著影响,较高或较低的水深都会影响挺水植物的地上和地下部分形态指标、相对生长速率指标和生理指标,而挺水植物通过调节形态和生理指标来适应和抵抗水深胁迫带来的不利影响.
b) 在高水深这一胁迫条件下,3种挺水植物都倾向于产生更多的总可溶性糖、SOD和MDA,显示出植物对高水深这一胁迫的生理适应性.
c) 在低水深条件下植物配置应优先配置较为不耐水淹的慈姑,高水深条件下优先配置较为耐水淹的菰,中水深条件下应以再力花为主. 考虑到再力花为外来种,目前尚未经过外来种入侵风险评估,在太湖沿岸带水生植物恢复中应慎重使用.
