新疆地区牛分枝杆菌29个毒力基因的突变分析
2021-11-04董玉慧罗力嘉李梦莹欧喜超刘春法范伟兴赵雁林周向梅
董玉慧 罗力嘉 李梦莹 欧喜超 刘春法 范伟兴 赵雁林* 周向梅*
(1.中国农业大学 动物医学院,北京 100193;2.宁夏大学 农学院,银川 750021;3.中国疾病预防控制中心,北京 102206;4.中国动物卫生与流行病学中心,山东 青岛 266032)
结核病是由毒力和传播能力不同的结核分枝杆菌复合群(Mycobacteriumtuberculosiscomplex,MTBC)成员感染人类和动物引起的一种人畜共患病[1]。MTBC成员主要包括引起人结核病的结核分枝杆菌、非洲分枝杆菌,以及从其他哺乳动物分离的牛分枝杆菌、田鼠分枝杆菌等[2-3]。牛结核病是一种主要由牛分枝杆菌引起的慢性消耗性疾病。牛分枝杆菌宿主范围广泛,可感染牛等多种动物及人类,给畜牧业生产带来重大的经济损失,并威胁人类健康,该病被世界动物卫生组织(OIE)列为B类动物疾病[4-5]。但是由于许多国家缺乏对牛结核病的长期监测和流行病学调查,牛分枝杆菌的流行率和对人类的影响被低估[6-7]。因此,评估牛分枝杆菌的遗传多样性及其与致病性和疾病严重程度的相关性,对养牛业的健康发展和公共卫生安全至关重要[7]。
MTBC可通过突变、重组和自然选择等途径实现全基因组多样化,从而在全球广泛流行和传播[8-9]。结核分枝杆菌系统发育谱系具有显著的地理分布特点,呈现出与人类宿主密切的共同进化趋势[9]。不同谱系的分离株因多种表型如耐药性、毒力和致病性而有所不同,这些表型影响疾病的严重程度,而多样性是由影响表达产物结构和功能的基因突变造成的[10]。利用转座子突变体库结合不同的体内筛选方法,鉴定出分枝杆菌大量的毒力基因,这些毒力基因大多数编码脂质代谢相关酶、细胞表面蛋白、调节因子和信号转导系统蛋白,影响分枝杆菌在宿主巨噬细胞内的存活[11]。目前,中国新疆地区流行牛分枝杆菌毒力变化和基因突变特征仍不明确。比对不同毒力菌株的毒力基因序列,找到关键突变位点,是发现细菌致病机理和进化规律的重要方法。Mycobacteriumbovis(M.bovis) C68004分离自上世纪60年代北京结核病牛,M.bovisN由本实验室近期从全身粟粒性结核病牛分离而来。在BALB/c小鼠模型上研究发现,与M.bovisC68004菌株相比,M.bovisN菌株表现出更强的致病性和更高的致死率。全基因组测序结果表明,M.bovisC68004菌株和M.bovisN菌株共存在750个SNPs (Single nucleotide polymorphisms)和145个INDELs(Insertion/Deletion)差异。与毒力因子数据库匹配后,发现29个毒力基因发生突变[12]。
本研究通过对分离自新疆地区的66株牛分枝杆菌的29个毒力基因进行测序,将其序列与标准株M.bovisAF2122/97及本实验室保存的M.bovisC68004和M.bovisN进行序列比对,初步评估新疆地区牛分枝杆菌的毒力情况,并进而探讨基因突变对牛分枝杆菌毒力的影响,为后续深入研究牛分枝杆菌的致病机理和进化规律奠定了基础。
1 材料与方法
1.1 试验材料
中性改良罗氏基础培养基(Celnovte,中国);细菌基因组DNA提取试剂盒(Tiangen,中国);Zihel & Neelsen染色试剂盒、PCR Mix、DL 2000 DNA Marker、6 x DNA loading Buffer(Solarbio,中国);PCR扩增仪和凝胶成像分析系统(Bio-Rad,美国);本实验中所用的试验菌株包括:从新疆地区分离的66株牛分枝杆菌(由中国疾病预防控制中心结核病预防控制中心保存),M.bovisC68004(购自中国兽医药品监察所),M.bovisN(由中国农业大学动物医学院国家动物传染性海绵状脑病实验室保存)。
1.2 试验方法
1.2.1牛分枝杆菌的培养
取200 μL冻存菌液,将其均匀涂于改良罗氏基础培养基上,37 ℃温箱培养2~3周。
1.2.2细菌基因组DNA的提取
在生物安全三级实验室(P3)中操作:刮取约20~100 mg(湿重)细菌新鲜培养物于1.5 mL螺旋口离心管中,水浴80 ℃灭活30 min,根据P3的操作规范要求,可将灭活的菌液移出P3,其他步骤参照细菌基因组DNA提取试剂盒说明书进行。
1.2.3引物的设计与合成
参考GenBank上已发表的29个牛分枝杆菌毒力基因序列,使用Primer 5.0软件设计引物序列,送至金唯智生物科技有限公司合成,引物的序列见表1。
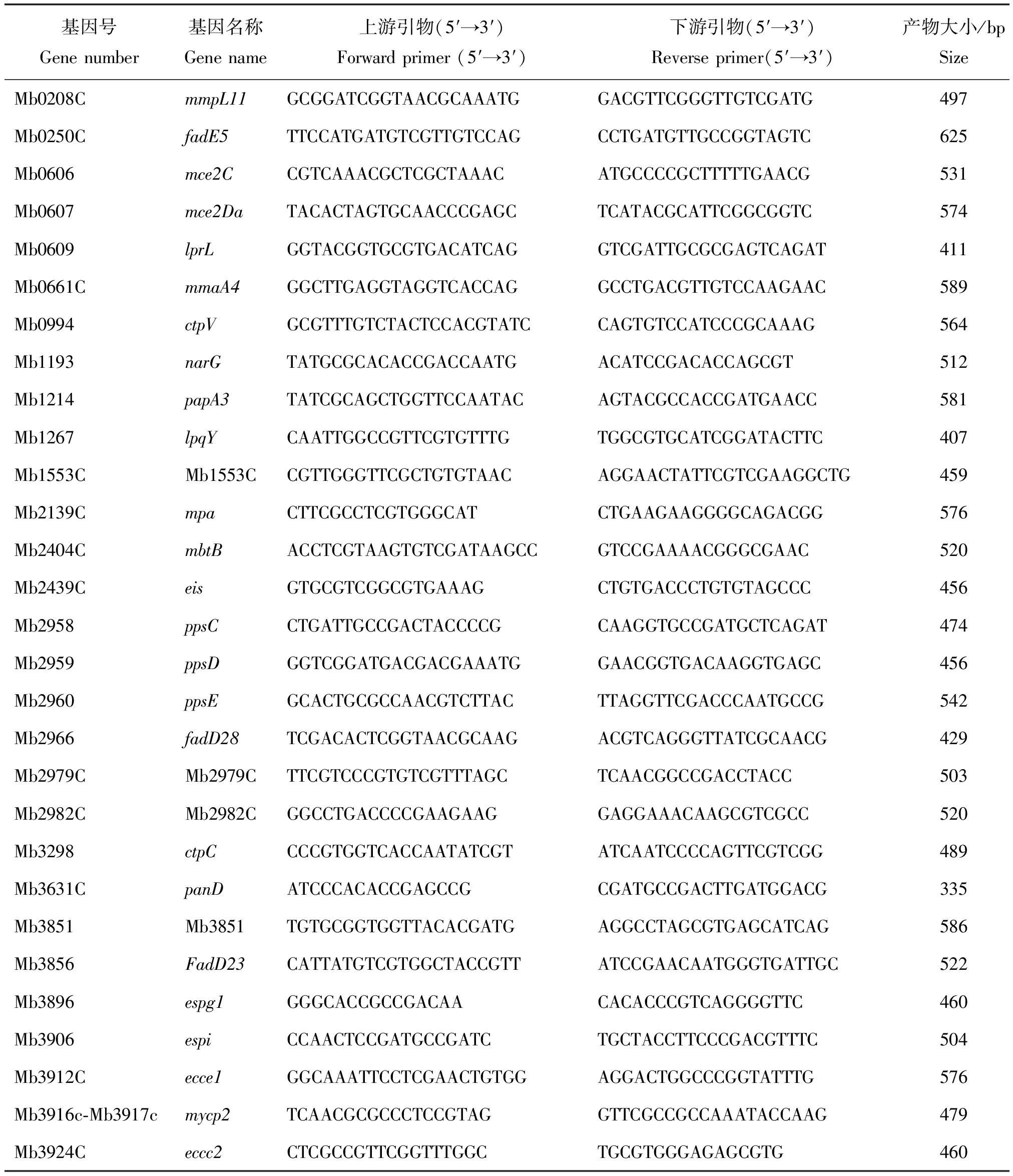
表1 新疆地区牛分枝杆菌29个毒力基因的引物序列和目的扩增产物大小Table 1 Primer sequences and targeted amplification product size of 29 virulence genes from M. bovis in Xinjiang
1.2.4PCR反应体系和程序
反应体系为25 μL,其中模板1 μL,PCR Mix 12.5 μL,ddH2O 10.5 μL,上游引物0.5 μL(10 μmol/L),下游引物0.5 μL(10 μmol/L)。反应程序为:95 ℃预变性20 min,95 ℃变性30 s,60 ℃退火30 s,72 ℃延伸60 s(35个循环),72 ℃延伸10 min,4 ℃保存。
1.2.5PCR产物回收和测序
将各基因的PCR产物送至金唯智生物科技有限公司进行Sanger测序,并在NCBI/GenBank上进行序列比对。
1.3 序列分析
利用PROVEAN和SOPMA在线分析软件预测基因突变对蛋白质功能的影响,评价牛分枝杆菌毒力基因突变对蛋白质功能或二级结构的影响。
PROVEAN软件通过执行BLAST搜索以识别同源序列,并生成PROVEAN得分,可预测单个或多个氨基酸替换、插入和删除对蛋白功能的影响。PROVEAN对单个氨基酸变异的功能预测效果与其他同类工具(如SIFT和PolyPhen-2)具有良好的一致性[13]。SOPMA是一种新的预测蛋白二级结构的方法,通过5种相互独立的方法进行预测,并将结果汇集整理成一个“一致预测结果”,具有较高预测蛋白二级结构的成功率[14]。
2 结果与分析
2.1 新疆地区牛分枝杆菌29个毒力基因的扩增
使用设计的引物,分别扩增66株牛分枝杆菌的29个毒力基因片段。PCR扩增产物经1.5%琼脂糖凝胶电泳分析,产物条带大小符合预期设计目的片段,随后进行Sanger测序(图1)。
M: DNA分子量标准Trans2K; 1~29: Mb0208C、Mb0250C、Mb0606、Mb0607、Mb0609、Mb0661C、Mb0994、Mb1193、Mb1214、Mb1267、Mb1553C、Mb2139C、Mb2404C、Mb2439C、Mb2958、Mb2959、Mb2960、Mb2966、Mb2979C、Mb3851、Mb3924C、Mb3916C-Mb3917C、Mb3896、Mb2982C、Mb3906、Mb3912C、Mb3298、Mb3631C、Mb3856.M: Trans2K DNA Marker; 1~29: Mb0208C、Mb0250C、Mb0606、Mb0607、Mb0609、Mb0661C、Mb0994、Mb1193、Mb1214、Mb1267、Mb1553C、Mb2139C、Mb2404C、Mb2439C、Mb2958、Mb2959、Mb2960、Mb2966、Mb2979C、Mb3851、Mb3924C、Mb3916C-Mb3917C、Mb3896、Mb2982C、Mb3906、Mb3912C、Mb3298、Mb3631C、Mb3856.图1 新疆地区牛分枝杆菌29个毒力基因扩增Fig.1 PCR amplification of 29 virulence genes of M. bovis in Xinjiang
2.2 新疆地区牛分枝杆菌临床分离株基因变异的检测
为研究牛分枝杆菌临床分离株毒力基因的变异情况,本研究将66株临床分离株与标准株M.bovisAF2122/97以及实验室保存的M.bovisC68004和M.bovisN的基因序列进行比对。结果发现,与标准株M.bovisAF2122/97基因序列相比,其中有17个毒力基因在超过50%的临床分离株存在突变(图2)。此外,在29个检测的毒力基因中,50%以上临床分离株与M.bovisN突变位点序列一致的共有25个基因,50%以上临床分离株与M.bovisC68004序列仅在3个基因中表现一致。这些结果表明中国新疆临床分离的牛分枝杆菌毒力基因突变特征与致病性更强的M.bovisN具有很高的相似性。
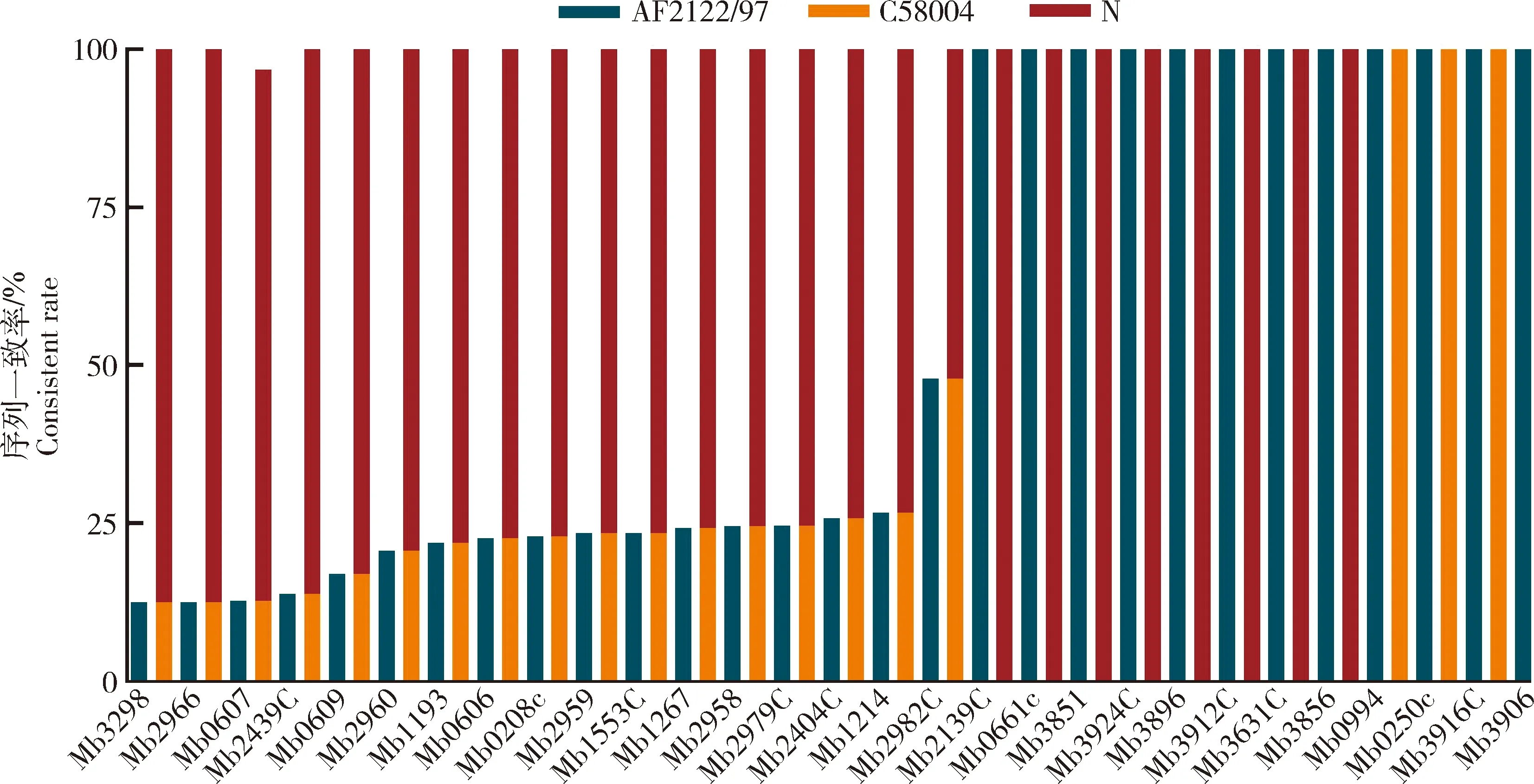
序列一致率:毒力基因突变位点与参比菌株序列一致的临床分离菌株数/临床总分离菌株数×100%。Consistent rate: Number of clinical isolates with the same virulence gene mutation site as the reference strain/umber of total cunleal isolates.图2 牛分枝杆菌临床分离株毒力基因与AF2122/97、C68004、N菌序列比较Fig.2 Comparison of virulence gene sequences in clinical isolates of M. bovis with AF2122/97,C68004 and N
2.3 基因变异对蛋白功能的影响
2.3.1SNP对蛋白功能的影响
蛋白质编码区的SNPs可影响氨基酸序列,从而改变蛋白质的结构和功能。在牛分枝杆菌临床分离株中的12个基因发现了广泛的碱基替换。为分析SNP对蛋白质功能的影响,本研究使用PROVEAN软件[13]预测非同义突变对蛋白生物学功能的影响,当PROVEAN评分等于或低于-2.5时,则该点突变对蛋白功能具有“有害”影响;当PROVEAN评分高于-2.5时,则为“中性”突变。结果揭示Mb2958、Mb2982C和Mb1553C基因的非同义突变PROVEAN评分均低于-2.5,表明可能对蛋白质生物学功能有显著影响。另外,其余基因的PROVEAN评分均高于-2.5,说明基因发生的这些突变对蛋白功能的影响不明显(图3)。
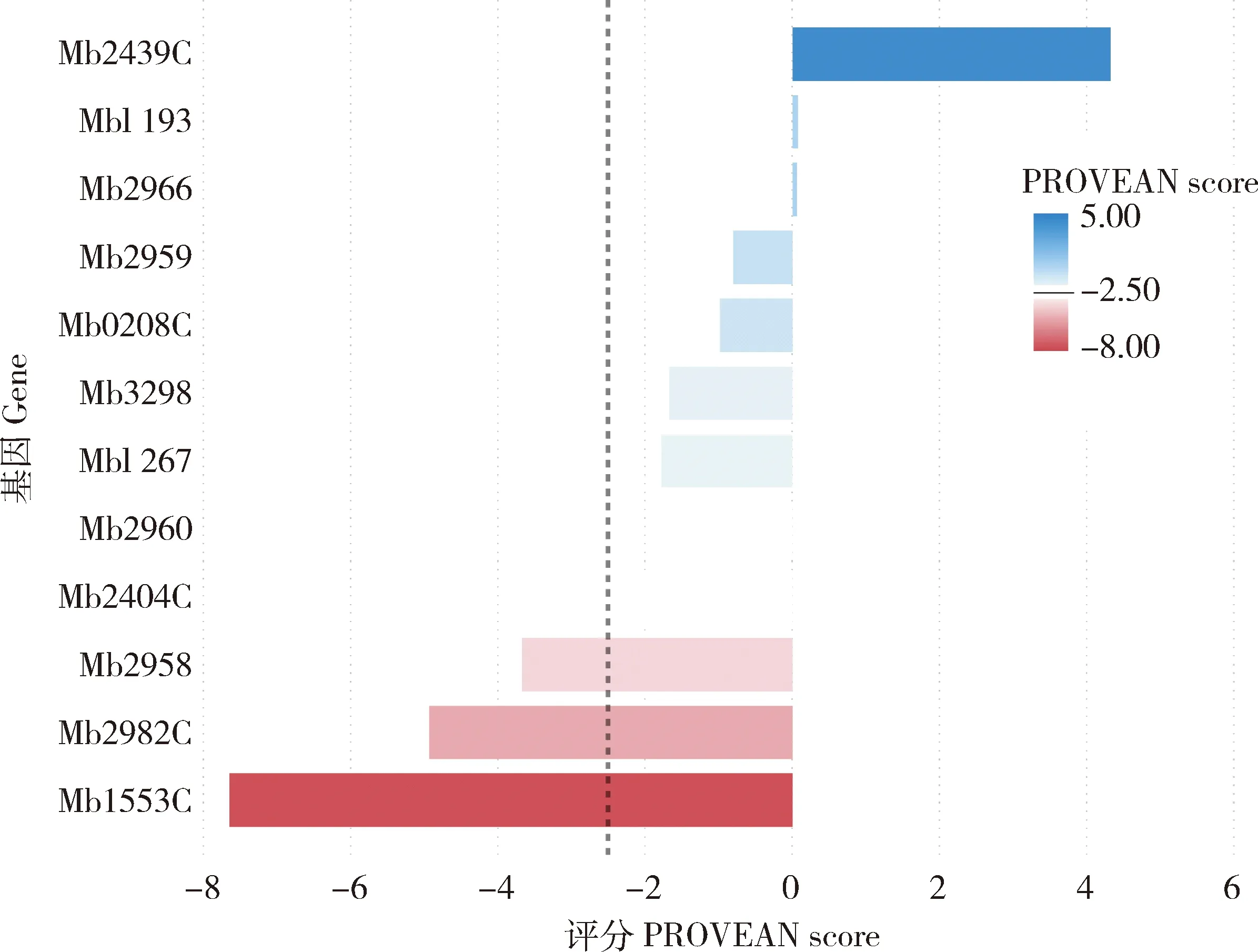
图3 预测牛分枝杆菌非同义SNP对蛋白质生物学功能的影响Fig.3 Prediction of the effect of nonsynonymous SNP in M.bovis isolates on the biological function of the proteins
2.3.2蛋白及其突变体二级结构的预测
使用SOPMA(Self-optimized prediction method with alignment)服务器[14]预测蛋白质及其突变体的二级结构。结果发现5种广泛存在于临床菌株的基因变异导致蛋白质的二级结构发生变化。Mb0607基因的第1 395位核苷酸插入一个碱基(C,胞嘧啶)导致肽链的延长(图4(a))。Mb0609基因编码的蛋白质终止密码子突变为Gln,使得翻译终止延后(图4(b))。Mb0606基因第195位氨基酸Gln突变为终止密码子,导致转录提前终止(图4(c))。Mb2979C基因的第232位核苷酸插入了一个碱基(C,胞嘧啶),这种移码突变造成了肽链的缩短(图4(d))。此外,Mb1214基因的第367位核苷酸插入一个鸟嘌呤,并引起下游的密码子发生改变,使肽链缩短(图4(e))。结果表明,这5种常见的基因变异可导致相应蛋白二级结构的改变。

W:野生型蛋白二级结构;M:突变体蛋白二级结构W: Secondary structure of wild-type protein; M: Secondary structure of mutant protein图4 牛分枝杆菌毒力因子基因突变对其蛋白二级结构影响的预测Fig.4 Prediction of effects of virulence gene mutants on the secondary structure of proteins
3 讨 论
由于全球多数地区缺乏对牛分枝杆菌基因组多样性的调查和可比的基因分型数据,目前,对牛分枝杆菌全球分布和进化的研究较少[4]。最近的一项研究表明牛分枝杆菌至少存在4个全球谱系,具有明显的地理分布特点[15]。但来自亚洲和非洲的牛分枝杆菌基因组数据匮乏,牛分枝杆菌全球分布谱系仍不完整[15-16]。本研究对我国新疆分离的66株牛分枝杆菌的毒力基因突变进行了初步研究,并将其毒力基因序列与两株牛分枝杆菌临床分离株(M.bovisC68004和M.bovisN)和牛分枝杆菌标准株M.bovisAF2122/97进行了序列比对。本团队之前研究发现,与M.bovisC68004株相比,M.bovisN株对小鼠的致病性很强,M.bovisN更强的致病性可能与其毒力基因的突变有关[12]。本研究在检测的29个毒力基因中,发现50%以上的临床分离株与M.bovisN突变位点序列一致的基因有25个。这在一定程度上反映了目前新疆地区流行牛分枝杆菌毒力的情况。
近年来,对结核分枝杆菌的致病性、毒力和持久性的分子基础研究取得了相当大的进展,牛分枝杆菌与结核分枝杆菌的基因组序列相似性超过99.95%[11,17],因此,牛分枝杆菌毒力基因的研究可参考结核分枝杆菌。本研究分析发现临床菌株广泛存在的Mb2958、Mb2982C和Mb1553C基因非同义突变对蛋白功能影响较大,对牛分枝杆菌毒力可能有一定影响。Mb2958(ppsC)与结核分枝杆菌Rv2933具有100%相似性,编码I型聚酮合成酶,催化脂类毒力因子Phthiocerol dimycocerosates(PDIM)和酚糖脂(Phenolic glycolipids,PGL)的生物合成关键步骤[18-19]。Mb2982C与结核分枝杆菌Rv2958C具有83.3%的相似性,编码参与PGL-tb合成的糖基转移酶。PGL包含有一个由长链β二醇结构形成的脂质核心,与多甲基分枝的脂肪酸形成二酯,存在于细胞壁,仅少量结核分枝杆菌可以合成PGL-tb,PGL-tb碳水化合物组分主要形式为2,3,4-三-O-甲基-L-吡喃岩藻糖基-α-(1-3)-L-吡喃鼠李糖基-α-(1-3)-2-O-甲基-L-吡喃鼠李糖[20-21]。而牛分枝杆菌由于基因移码突变合成一种截短形式的PGL-tb,称为分枝杆菌糖脂B,碳水化合物区域限制为2-O-甲基鼠李糖[22]。PGL是致病性分枝杆菌的一个重要毒力因子,在抗氧化应激、免疫调节和细胞趋化过程中发挥作用,该化合物有多种结构变异。这些结构改变的影响虽尚未被评估,但可能解释分枝杆菌引起疾病的不同特征[20]。糖肽脂(Glycopeptidolipids,GPLs)是存在于许多分枝杆菌表面的特异性糖脂类,其特征是糖基化模式的高度变异性,具有多种生物活性,Gtf3是一种催化合成三糖基化GPLs的糖基转移酶[23]。Mb1553C与结核分枝杆菌Rv1526C基因序列具有100%一致性,为Gtf3的同源物。Mb2958、Mb2982C和Mb1553C基因突变均可能导致其编码的酶活性发生改变,从而对细胞壁脂类合成产生影响,导致牛分枝杆菌致病性增强。
本研究发现超过半数临床菌株存在的Mb0607(Mce2Da)、Mb0609(lprL,Mce2E)、Mb0606(Mce2C)、Mb2979C和Mb1214(Papa3)基因突变导致了蛋白二级结构的显著变化。哺乳动物细胞入侵(Mammalian cell entry,Mce)基因是Mce操纵子的组成部分,其编码的蛋白在分支杆菌进入哺乳动物细胞和在巨噬细胞存活过程中发挥有重要作用,哺乳动物细胞入侵蛋白在分枝杆菌细胞表面的位置与Mce操纵子在哺乳动物细胞侵袭中的潜在作用一致,因此被视为重要的毒力因子[24-25]。本研究表明基因突变导致Mce2Da、Mce2E和Mce2C蛋白二级结构发生显著变化,其中,Mce2E发生于终止密码子区域的基因突变,使其表达产物和M.tuberculosisH37Rv具有100%一致性,因此进一步对这些Mce操纵子进行深入研究将有助于阐明牛分枝杆菌毒力变异的机制。Mb2979C与结核分枝杆菌Rv2955C相似性为99.7%,Rv2955C参与结核分枝杆菌细胞壁PGL-tb的生物合成[26]。Mb1214与结核分枝杆菌Rv1182序列100%一致,编码Polyacyltrehalose(PAT)生物合成所必需的酰基转移酶[27]。后2种基因突变导致的编码蛋白结构的变化可能会对其生物效应产生影响,从而提高牛分枝杆菌的毒力。
基因研究为分枝杆菌属的遗传多样性提供了一个精细化的理解,这些结果为MTBC的宏观和微观进化提供了一个新的视角[28-29]。本研究在Mb2958、Mb2982C、Mb1553C、Mb0607、Mb0609、Mb0606、Mb2979C和Mb1214这8个毒力基因中发现了新疆牛分枝杆菌临床分离株广泛存在的突变,这些基因突变可能与牛分枝杆菌在牛群中更强的适应性和致病性有关,其具体机制尚需进一步研究。
综上,本研究通过对牛分枝杆菌毒力基因进行检测、测序和比对,并分析基因突变对其编码蛋白功能和结构的可能影响,初步反映了我国新疆地区牛分枝杆菌流行菌株的毒力情况,为后续研究牛分枝杆菌分子流行病学和阐明牛分枝杆菌致病机理及进化规律奠定了基础。
