氮添加对不同植被类型中毛竹与常绿阔叶林木生长的影响
2021-09-12赖雁南钟全林李宝银程栋梁余华徐朝斌薛翔海郑文婷
赖雁南 钟全林 李宝银 程栋梁 余华 徐朝斌 薛翔海 郑文婷



摘 要 氮沉降背景下毛竹向常綠阔叶林扩张问题是当前我国亚热带区域面临的主要生态问题与研究热点之一。以毛竹、竹阔混交林及常绿阔叶林为对象,通过1 a的氮添加试验,分析毛竹与常绿阔叶林木在不同植被类型中生长的差异,以探究氮沉降对不同植被类型中毛竹与常绿阔叶林木生长的影响。结果显示:(1)竹阔混交林中毛竹的平均胸径增长量、平均单株生物量增长量及生物量相对生长速率高于毛竹林中的毛竹,但竹阔混交林中阔叶林木的平均胸径增长量、平均单株生物量增长量及生物量相对生长速率则显著低于常绿阔叶林中的阔叶林木(P<0.05);(2)低氮添加会减小毛竹在竹阔混交林与毛竹林中的个体差异,但会增大常绿阔叶林木在竹阔混交林与常绿阔叶林之间的生长差距,低氮添加有利于毛竹向常绿阔叶林扩张;(3)高氮添加会显著抑制毛竹的生长,一定程度上可减缓毛竹向常绿阔叶林扩张的速度。
关键词 氮添加;生物量;相对生长速率;毛竹;常绿阔叶林木
中图分类号:S714.8;S718.54 文献标识码:A doi:10.13601/j.issn.1005-5215.2021.02.001
Abstract The expansion of Phyllostachys pubescens to evergreen broadleaved forests under the background of nitrogen deposition is one of the main ecological problems and research hotspots facing China's subtropical regions. Taking Phyllostachys edulis,Phyllostachys edulisevergreen broadleaved mixed forest and evergreen broadleaved forest as the object,the growth difference between Phyllostachys edulis & evergreen broadleaved forest in different vegetation types were analyzed through 1year nitrogen addition test,and explores the influence of Phyllostachys edulis expansion into evergreen broadleaved forest under the background of nitrogen deposition on the growth of Phyllostachys edulis and evergreen broadleaved forest. Results showed that: (1) In Phyllostachys edulisevergreen broadleaved mixed forest average growth in DBH,average individual biomass growth and biomass per plant relative growth rate is greater than Phyllostachys edulis in Phyllostachys edulis forest,Phyllostachys edulisevergreen broadleaved mixed forest,evergreen broadleaved forest significant differences mainly growth parameters,Phyllostachys edulis expansion leads to the average individual DBH growth rates of evergreen broadleaved forest biomass growth and biomass per plant and average relative growth rate dropped significantly;(2) Low nitrogen addition can reduce the growth difference of Phyllostachys edulis in Phyllostachys edulisevergreen broadleaved mixed forest and Phyllostachys edulis forest,but increase the growth difference of evergreen broadleaved forest in Phyllostachys edulisevergreen broadleaved mixed forest and evergreen broadleaved forest. Low nitrogen addition is beneficial to the expansion of Phyllostachys edulis into evergreen broadleaved forest;(3) The addition of high nitrogen can significantly inhibit the growth of Phyllostachys edulis and alleviate the expansion of Phyllostachys edulis to evergreen broadleaved forest.
Key words nitrogen addition;biomass;relative growth rate;Phyllostachys edulis;evergreen broad leaved trees
氮(N)是陆地生态系统净初级生产力最主要的限制元素之一[1,2]。工业革命以来,人类生产生活方式的改变使得全球大气活性氮化合物含量上升 [3],导致大气氮沉降格局的改变。当前世界氮沉降分布中心正由欧美等老工业基地转移至各发展中国家,出现由温带逐渐扩散至热带亚热带的趋势[4]。随着经济快速发展,我国部分地区已出现高氮沉降问题,特别是在南方亚热带森林区域,氮沉降量已接近欧洲最高氮沉降水平[5-7]。预计未来几十年我国氮沉降量仍将持续增加,日益严峻的氮沉降问题已成为我国生态建设中被关注的热点[8]。
氮沉降对植物及森林生态系统的影响受其沉降量的不同而表现不同 [9],适度的氮沉降能增加土壤可利用氮含量,促进植物生长与生物量的积累,提高生态系统的生产力,但过量的氮沉降则会抑制植物的生长,降低生态系统的生物多样性[10]。氮沉降对植物生长的影响还取决于树种差异,不同树种对氮沉降的响应机制不同,导致其对氮的利用和配置存在差异[11]。有研究发现,氮添加能够提高土壤中活性N含量,从而促进植物生长,提高草原生态系统总生物量 [12];氮添加量与植物地上生物量呈显著正相关 [13],但也有人研究发现,氮添加量与生物量呈负相关关系,较低的氮添加可使得中国东北落叶阔叶林的生物量达到最大值[14]。林木对氮添加的响应程度,不仅受到氮添加量的影响,还与研究区域、树种及施氮量、氮肥种类、施氮年限等因素有关[15-18]。目前,国内外在氮添加对森林生态系统方面的研究主要集中于同一生态系统中森林的生长状况、土壤理化性质、土壤微生物及植物凋落物分解等方面[19],而有关氮添加对植物在不同森林类型中的生物量及其相对生长速率差异的影响研究则相对较少。因此有必要开展这方面研究,以探究毛竹及常绿阔叶树种在不同林分类型内对氮添加的响应特征,为进一步开展氮沉降背景下常绿阔叶林可持续经营技术及揭示氮沉降背景下毛竹向常绿阔叶林内的扩张机制等提供理论依据。
常绿阔叶林为我国亚热带地区的典型地带性植被,其物种多样性丰富,在保护区域生态环境、平衡全球性碳水循环及维持人类社会可持续发展等方面发挥着重要作用[20-22]。毛竹(Phyllostachys edulis)是禾本科刚竹属的一种散生乔木状竹类植物。常绿阔叶林周边的毛竹依靠地下根茎(即竹鞭)向常绿阔叶林区蔓延而实现种群扩张,致使阔叶纯林演替为竹阔混交林,甚至在人为干预下逐渐形成毛竹林[23]。毛竹扩张导致森林生态系统的功能与景观质量退化问题正逐渐为林学及生态学等学科相关学者所关注。已有研究发现,毛竹扩张引发了诸多生态环境问题,但人们往往因其具有较高的经济效益而忽视其造成的负面影响[24,25]。因此,研究氮添加对毛竹与常绿阔叶树种生长的影响,不仅有助于揭示毛竹与常绿阔叶树种对大气氮沉降的响应机制,对开展常绿阔叶林保护技术研究等也具有重要现实指导意义。
1 研究区概况与试验方法
1.1 研究区概况
试验地位于江西阳际峰自然保护区,地处江西省东北部,武夷山北麓(27°51′10″—28°02′20″ N, 117°11′30″—117°28′40″ E)。保護区森林总面积10 946 hm2,其中活立木蓄积量622 802 m3,立竹量9.8万株,森林覆盖率高达99.7%,天然常绿阔叶植被类型分布面积占总面积的68%,为我国亚热带常绿阔叶林的典型代表,同时也是毛竹适生区[26]。该地属中亚热带湿润季风气候区,年均温11.4~18.5 ℃,年均降水量1 870~2 191.3 mm,年均无霜期200~256 d,四季分明,温暖湿润,水热条件良好,有利于动植物生长发育,为试验研究提供了理想的平台。
1.2 模拟氮沉降试验设计
于2017年3—4月分别在天然毛竹林、天然常绿阔叶林及其中间过渡地带的竹阔混交林内各设置9个样地,共27个样地,其中毛竹林样地规格为10 m×10 m,常绿阔叶林与竹阔混交林样地规格均为15 m×15 m;3种植被类型均为经自然更新达20 a以上的天然林,所选设的样地海拔约为730 m;同一森林类型样地海拔相近,毛竹林海拔相对低于常绿阔叶林。分别植被类型在所选设的9个样地中随机设置对照(CK,0 kg a-1)3种处理,每处理设置3个重复。所添加的氮肥为尿素,各处理的氮添加量中均未包含自然氮沉降量。将所确定的年氮添加量均分成各月添加量,并换算成各样地面积的月氮添加量。于2017年5月—2018年8月,于每月月底晴朗无风的9:00—11:00进行,持续施氮16个月。每次喷施之前,将称取的氮肥溶解于蒸馏水中(毛竹林样方是将氮肥溶解于10 L水中,竹阔混交林与天然常绿阔叶林样方是将氮肥溶解于20 L水中),使用双喷头喷雾器将其水溶液人工来回均匀喷洒在林地上,其中,对不施肥的对照组则喷洒等量的蒸馏水,以消除误差。3种植被类型样地基本信息见表1。
1.3 指标选取及测定
分别于2017年4月第1次施肥之前及2018年8月对样地内毛竹和胸径≥5.0 cm的林木进行调查,测定胸径、株高(常绿阔叶树种株高为树冠最高处离地面的垂直距离,毛竹的为枝下高)及树龄,用于计算各样地植物的生物量。
1.3.1 常绿阔叶林木与毛竹平均胸径与胸径增长量的计算
常绿阔叶林木与毛竹平均胸径(DBH)计算公式如下:
DBH=1N∑ni=1(DBHi)2 (1)
式中:DBH为常绿阔叶林木(或毛竹)的平均胸径(cm);n为样地内胸径≥5.0 cm常绿阔叶林木(或所有毛竹)的数量;DBHi为样地内第i株常绿阔叶林木(或毛竹)的胸径。
常绿阔叶林木与毛竹胸径增长量计算公式如下:
ΔDBH= DBH2- DBH1(2)
式中:ΔDBH为常绿阔叶林木(或毛竹)的平均胸径增长量(cm);DBH1与DBH2分别为期初与期末常绿阔叶林木(或毛竹)的平均胸径(cm)。
1.3.2 常绿阔叶林木与毛竹生物量的计算
(1)常绿阔叶林木生物量的计算采用材积源生物量法[27]和已有文献所建立的回归方程[28]计算不同植被类型常绿阔叶林木的生物量。回归方程如下:
1.3.3 生物量增长量与相对生长速率的计算
1.4 数据处理
利用Microsoft Excel 2010软件对数据预处理。运用SPSS 20.0软件对氮添加和植被类型影响下单株毛竹及阔叶林木生长指标进行双因素方差分析;对毛竹和林木在同一植被类型不同氮添加水平下及同一氮添加水平下同一树种在不同植被类型中的生长指标进行单因素方差分析,用最小显著差异法(LSD) 进行处理间差异显著性比较(α=0.05),使用Origin2018软件进行绘图。
2 结果与分析
2.1 植被类型与氮添加对毛竹与常绿阔叶林木生长的影响
雙因素方差分析结果(表2)显示,生长在毛竹纯林与竹阔混交林中的毛竹仅其平均单株生物量存在显著差异(P<0.05),但生长在常绿阔叶林与竹阔混交林中的常绿阔叶林木的平均高、平均单株胸径、平均单株生物量、平均单株生物量相对生长速率、单位面积生物量相对生长速率5个指标均有显著影响(P<0.05);氮添加对毛竹各生长指标均无显著影响,但对林木的生物量及生物量相对生长速率则有极显著影响(P<0.01)。植被类型和氮的交互作用对毛竹没有显著影响,但对阔叶林木的平均单株生物量和平均单株生物量相对生长速率有极显著影响(P<0.01)。
2.2 毛竹与常绿阔叶林木在不同植被类型中的生长差异
通过对未添加氮的毛竹林、常绿阔叶林及竹阔混交林数据进行分析,结果发现,生长在竹阔混交林内的毛竹,其平均单株生物量增长量及生物量相对生长速率虽高于其在毛竹林内的生长量,但两者差异不明显(P>0.05,图1A),表明生长在竹阔混交林中的毛竹个体生物增长量虽大于毛竹纯林中的个体生物增长量,但两者差异不明显;生长在竹阔混交林内的常绿阔叶林木,其平均单株生物增长量及平均单株生物量相对生长速率明显低于其生长在常绿阔叶林内(P<0.05,图1B),说明与竹阔混交林相比,常绿阔叶林木更适合在常绿阔叶林环境中生长。
2.3 氮添加对毛竹林及竹阔混交林中毛竹生长的影响
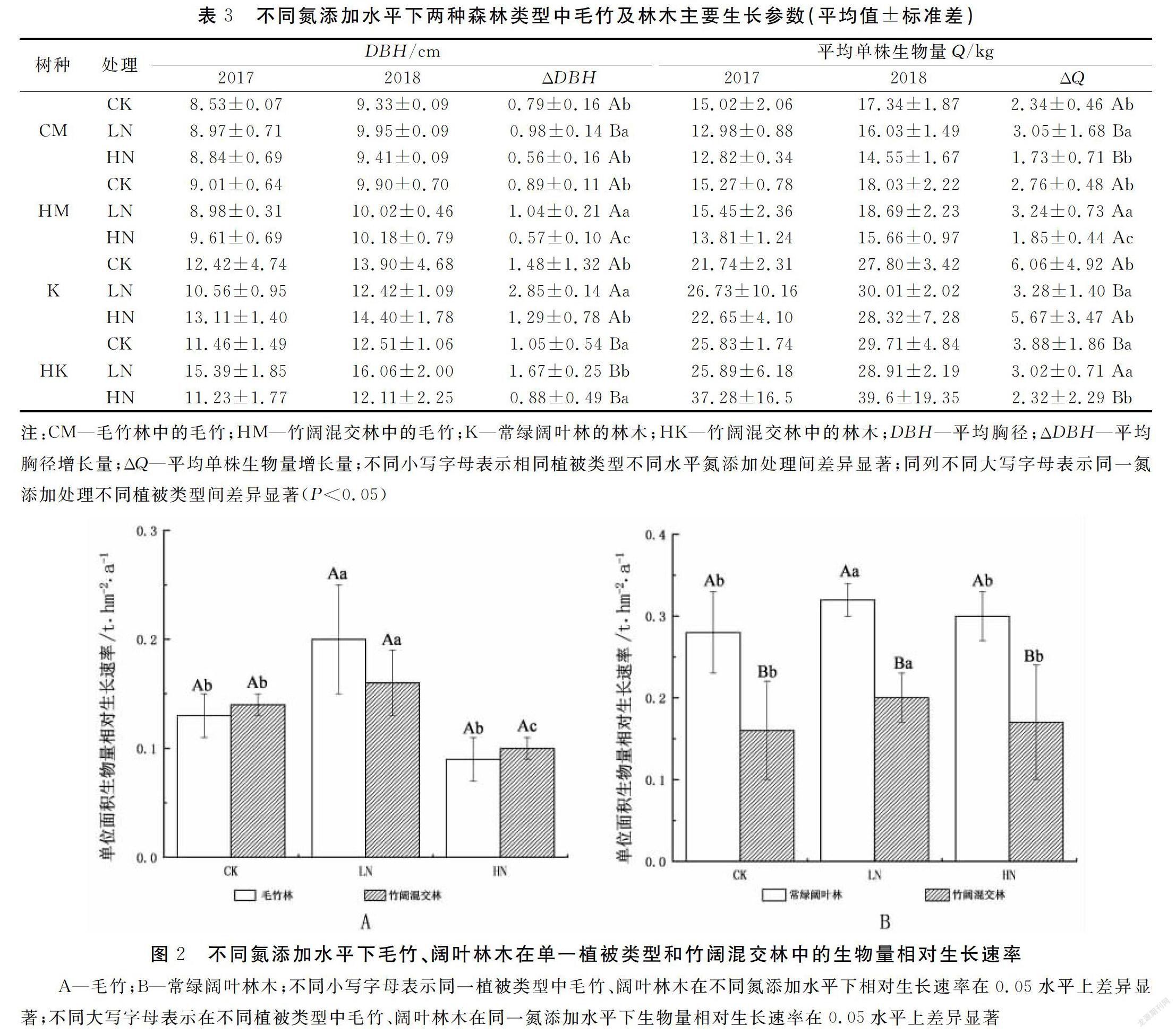
由表3和图2A可以看出,毛竹林中的毛竹在添加低氮后,平均单株胸径增长量、平均单株生物量增长量及生物量相对生长速率均有显著增加(P<0.05),但高氮添加则会造成毛竹林中的毛竹平均单株胸径增长量、平均单株生物量增长量及生物量相对生长速率均有减少,且与对照组差异不明显(P>0.05)。
在竹阔混交林中,氮添加会显著影响竹阔混交林中毛竹的平均单株胸径增长量、平均单株生物量增长量及生物量相对生长速率。低水平的氮添加,会显著增大竹阔混交林中毛竹的平均单株胸径增长量、平均单株生物量增长量、生物量相对生长速率(P<0.05);高水平的氮添加,则会显著减少竹阔混交林中毛竹的平均单株胸径增长量、平均单株生物量增长量、生物量相对生长速率(P<0.05)(表3)。
另从表3与图2A中可看出,氮添加会在一定程度促进竹阔混交林中的毛竹生长,使混交林中的毛竹胸径生长量、平均单株生物生长量及单位面积生物量生长速率高于毛竹纯林,但差异不明显,说明氮添加背景下有利于毛竹向竹阔混交林扩张。
2.4 氮添加对常绿阔叶林及竹阔混交林中林木生长的影响
由表3和图2B可以看出,在常绿阔叶林中的阔叶林木在添加低氮后,平均单株胸径增长量、平均单株生物量增长量、生物量相对生长速率均有显著增加(P<0.05)。高氮添加后,阔叶林木的平均单株胸径增长量、平均单株生物量增长量及生物量相对生长速率无显著变化。
在竹阔混交林中,不同水平氮添加对混交林中的阔叶林木平均单株胸径增长量、平均单株生物量增长量、生物量相对生长速率影响程度不同。低水平氮添加,会显著增大竹阔混交林中阔叶林木的平均胸径增长量、平均单株生物量增长量、生物量相对生长速率(P<0.05);但高氮添加则对竹阔混交林中阔叶林木的平均胸径增长量、平均单株生物量增长量、生物量相对生长速率影响不明显。
上述结果表明,不施氮时毛竹扩张会显著抑制林木的生长;低氮添加会显著促进常绿阔叶林中的阔叶林木生长,会增大阔叶林木在常绿阔叶林和竹阔混交林中生长的差异,有利于毛竹向阔叶林扩张;高氮添加对竹阔混交林中的毛竹和阔叶林都有抑制作用,毛竹受抑制程度强于阔叶林,因此高氮一定程度上缓解了毛竹向阔叶林扩张的速率。
3 讨论
3.1 毛竹与常绿阔叶林木在不同植被类型中的生长差异
常绿阔叶植被类型分布区同时也是毛竹适生区,毛竹扩张引发森林景观变化、阔叶树种生长减缓及碳储量的改变[30]等生态变化,这些变化可表现为植物部分形态特征和生长指标的改变。本研究中混交林中的毛竹平均胸径增长量、枝下高、平均单株生物量增长量相比毛竹林均有增加但不显著,毛竹单位面积生物量在混交林和毛竹林存在显著差异。这一结论符合毛竹的生长特性,春笋破土前竹鞭的发育、土壤状况、水肥等条件决定了春笋的大小,春笋后期不会增大决定了新竹胸径的大小[31,32]。
种间竞争是塑造植物形态和生活史的主要动力之一,Grime理论认为,相对生长速率(RGR)是衡量植物竞争的关键指标之一[33,34]。本文通过生物量RGR的分析发现,受毛竹扩张影响形成的竹阔混交林中常绿阔叶林木RGR有显著下降,并且竹阔混交林中阔叶林木的平均胸径增长量、单株生物量增长量、单位面积生物量相比常绿阔叶林也均有显著下降,这与赵雨虹的研究结果[35]相符,表明随毛竹扩张程度加剧,常绿阔叶树胸径、生长速率逐渐降低。这很大程度可能缘于毛竹具有生长快速、繁殖力强、形态多样、集团协同等内禀优势[36]。毛竹这些生长特点有利于毛竹在常绿阔叶林中的扩张,毛竹扩张后在竹阔混交林中相对生长速率提高,而毛竹和阔叶林木的种间竞争导致常绿阔叶林木的相对生长速率下降,各项生长指标增长量减少。
毛竹对常绿阔叶林的扩张影响了常绿阔叶林的系统结构和功能。森林植被生物量是研究森林生态系统结构与功能的重要指标,生物量估算已经成为研究森林结构和功能的重要内容。本研究通过计算毛竹及常绿阔叶林木在竹阔混交林与毛竹林、常绿阔叶林中平均单株生物量增长量的差异,发现毛竹在竹阔混交林(2.76 kg -1)中平均单株生物量增长量(ΔQ)与在毛竹林(2.34 kg -1)相比提高17.9%,而常绿阔叶林木在竹阔混交林中单位面积生物量增长量(3.88 kg -1)与在常绿阔叶林(6.06 kg -1)相比下降超过36.0%。这是因为当植物入侵原有森林生态系统时,会对其碳循环产生影响,进而导致植被生物量变化[37],如杨清培等[38]在研究毛竹入侵常绿阔叶林生态系统时发现植被碳储量出现显著降低,森林的总碳储量减少了15.37%, 其中植被碳储量减少了12.01%, 土壤碳储量减少了17.30%。毛竹擴张使得在竹阔混交林中毛竹生长加快,常绿阔叶林木生长减慢,从而减少常绿阔叶林物种多样性及森林植被碳储量,影响了常绿阔叶林生态系统的稳定性。
3.2 不同氮添加对毛竹及常绿阔叶林木生长的影响
氮沉降会改变森林的物种生物量的增长速率,其变化的程度取决于森林种类、林木径级与氮沉降浓度等因素的综合影响[38]。本研究中低浓度氮添加促进毛竹生长,高浓度氮添加抑制毛竹生长,表现为毛竹的平均单株生物量增长量、生物量相对生长速率随氮添加量增加(50、100 kg hm-2 -1)出现先升高后下降的变化,这和杜莹等[39]研究发现随添加的氮增加(30、60、90 kg -2 -1)表现为先升高后下降结论相符。这可能是因为不同水平的氮添加对毛竹光合作用的影响不同,进而影响了毛竹生物量的变化。刘宁的研究结果也支持了这一猜想,氮沉降促进了毛竹的光合速率,但是长期高水平的氮沉降会对毛竹的光系统Ⅱ产生一定伤害[40]。氮添加缩小了毛竹在毛竹林和竹阔混交林中生长的差异。这可能和单位面积内物种间竞争及环境最大承载量有关,毛竹林的毛竹在单一植被类型中生长不存在种间竞争,限制样地中毛竹最大生长数量的主要是环境最大承载量而不是氮素含量的高低。竹阔混交林中,毛竹与阔叶林竞争生长,毛竹对氮素的更高效利用有利于毛竹侵占阔叶树种更大的生长空间,最终表现出氮添加缩小了毛竹在毛竹林和竹阔混交林中的生长差异,都趋向于样地毛竹的最大环境承载量。
中国东北的落叶阔叶林的大量研究得出较低的氮添加(25 kg hm-2 a-1)导致最大的净生产力,随着氮添加的增加(50 kg hm-2 a-1)正面效应降低,氮添加量最大时(75 kg -2 -1)正面效应消失[41]。本研究中低氮添加会显著增加常绿阔叶林木的平均胸径增长量、平均单株生物量增长量及生物量相对生长速率,高氮添加对阔叶林木的生长无显著抑制作用。这可能和试验林下喷施氮肥的施肥方式有关。尿素适宜深施覆土,尿素在转化前是分子态的,不能被土壤吸附,容易随水流失,转化后的氨也易挥发。而高氮仍对竹阔混交林中的毛竹表现出显著抑制作用,这可能和毛竹的氮饱和值低于阔叶林木有关,陶晨悦等的研究支持这一猜想,干旱胁迫条件下,随着氮沉降水平的增加,毛竹生物量逐渐增大,但当氮沉降量大于60 kg hm-2 a-1时,生物量不再增大甚至有减小的趋势[42]。这表明氮素添加对毛竹的正面效应在氮浓度超过60 kg hm-2 a-1时消失,阔叶林木的氮饱和值要高于毛竹。低氮添加会加大阔叶林木在常绿阔叶林和竹阔混交林中生长的差异。这也是在竹阔混交林中物种间竞争能力低于毛竹的表现,短期的低氮添加有利于毛竹向常绿阔叶林的扩张。高水平的氮添加会显著抑制竹阔混交林中毛竹生长,对阔叶林木抑制作用不显著,表明短期的高氮添加一定程度上能缓解毛竹在常绿阔叶林中的扩张趋势。
上述分析结果仅是16个月的氮添加处理试验结果,随着持续的氮沉降,可能有些结果会发生变化。另外,毛竹扩张的地下根系入侵是扩张过程中非常关键和重要的部分,本研究受限于试验条件,没有探讨毛竹和林木地下部分对毛竹扩张和氮添加的响应。今后还需对其作进一步的深入研究,有利于进一步揭示毛竹在氮沉降背景下对阔叶林入侵的机制。
4 结论
毛竹扩张后,竹阔混交林中毛竹的平均单株生物量增长量及生物量相对生长速率均大于毛竹林,竹阔混交林中的林木平均单株胸径增长量、平均单株生物量增长量及相对生长速率相比常绿阔叶林有显著下降,表明毛竹扩张能够在一定程度上促进毛竹生长,显著抑制阔叶林木的生长;低氮添加会加大阔叶林木在竹阔混交林和常绿阔叶林的生长差异,减小毛竹在竹阔混交林和毛竹林的生长差异,有利于毛竹向常绿阔叶林的扩张;高氮添加会显著抑制毛竹生长,对林木生长无显著抑制作用,一定程度上可缓解毛竹向常绿阔叶林扩张。
参考文献:
[1] 周晓兵,张元明.干旱半干旱区氮沉降生态效应研究进展[J].生态学报,2009,29(7):3835-3845
[2] David S,Kathleen K.Nitrogen Limitation of Net Primary Productivity in Terrestrial Ecosystems Is Globally Distributed[J].Ecological Society of America,2008,89(2):371-379
[3] Galloway J,Dentener F,Capone D, et al.Nitrogen Cycles: Past,Present,and Future[J].Kluwer Academic Publishers,2004,70(2):153-226
[4] 鲁显楷,莫江明,张炜,等.模拟大气氮沉降对中国森林生态系统影响的研究进展[J].热带亚热带植物学报,2019,27(5):500-522
[5] 王小治,朱建国,高人,等.太湖地区氮素湿沉降动态及生态学意义:以常熟生态站为例[J].应用生态学报,2004(9):1616-1620
[6] 孙本华,胡正义,吕家珑,等.大气氮沉降对阔叶林红壤淋溶水化学模拟研究[J].生态学报,2006(6):1872-1881
[7] MacDonald J A,Dise N B,Matzner E,et al.Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests[J].Global Change Biology,2002,8(10):1028-1033
[8] 常运华,刘学军,李凯辉,等.大气氮沉降研究进展[J].干旱区研究,2012,29(6):972-979
[9] 张歆阳,王乔姝怡,邢娟,等.氮添加对武夷山米槠径向生长的影响[J].北京大学学报:自然科学版,2017,53(6):1143-1149
[10] 邹宇星,钟全林,游雅玲,等.短期氮-水处理对刨花楠幼苗细根根序形态的影响[J].应用生态学报,2018,29(7):2323-2329
[11] Hyvnen R,Persson T,Andersson S,et al.Impact of longterm nitrogen addition on carbon stocks in trees and soils in northern Europe[J].Biogeochemistry,2008,89(1):121-137
[12] 潘庆民,白永飞,韩兴国,等.氮素对内蒙古典型草原羊草种群的影响[J].植物生态学报,2005(2):311-317
[13] 李祿军,曾德慧,于占源,等.氮素添加对科尔沁沙质草地物种多样性和生产力的影响[J].应用生态学报,2009,20(8):1838-1844
[14] Yan G,Xing Y,Wang J,et al.Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China:Effects of nitrogen deposition[J].Agricultural & Forest Meteorology,2018,248:70-81
[15] 汪金松.模拟氮沉降对暖温带油松林土壤碳循环过程的影响[D].北京:北京林业大学,2013
[16] Gundersen P,Emmett B A,Kjonaas O J,et al. Impact of nitrogen deposition on nitrogen cycling in forests:A synthesis of NITREX data[J].Forest Ecology and management,1998,101(1):37-55
[17] 周利勋,刘广平,王金波.落叶松人工林的施肥效应[J].东北林业大学学报,2004(2):16-18
[18] RappeGeorge M O,Gardenas A l,Kleja D B.The impact of four decades of annual nitrogen addition on dissolved organic matter in a boreal forest soil[J].Biogeosciences Discussions,2012,10(3):1365-1377
[19] 于美佳,叶彦辉,韩艳英,等.氮沉降对森林生态系统影响的研究进展[J].安徽农业科学,2021,49(3):19-24,27
[20] 李超.中亚热带九种常绿阔叶树种幼苗叶结构型性状相关性与叶经济谱分析[D].北京:中国林业科学研究院,2016
[21] 周博,范泽鑫,杞金华.哀牢山中山湿性常绿阔叶林木径向生长季节动态及其对气候因子的响应[J].生态学报,2020,40(5):1699-1708
[22] Xu Y,Zhang Y,Yang J,et al.Influence of tree functional diversity and stand environment on fine root biomass and necromass in four types of evergreen broad-leaved forests[J].Global Ecology and Conservation,2020,21(C):e00832-e00832
[23] 《江西森林》编委会.江西森林[M].南昌:江西科学技术出版社,1986:220-230
[24] 欧阳明,杨清培,陈昕,等.毛竹扩张对次生常绿阔叶林物种组成、结构与多样性的影响[J].生物多样性,2016,24(6):649-657
[25] 童冉,周本智,姜丽娜,等.毛竹入侵对森林植物和土壤的影响研究进展[J].生态学报,2019,39(11):3808-3815
[26] 乐新贵.阳际峰自然保护区生态经济评价[D].南京:南京林业大学,2005
[27] 曹军,张镱锂,刘燕华.近20年海南岛森林生态系统碳储量变化[J].地理研究,2002(5):551-560
[28] Fang J Y, Chen A P, Peng C H, et al.Changes in forest biomass carbon storage in China between 1949 and 1998[J].Science,2001,292(5525):2320-2322
[29] 曾掌权, 田育新, 戴成栋, 等.湖南毛竹林生物量模型研究[J].湖南林业科技,2016,43(6):56-59
[30] 蔡爱明. 生物和非生物压力对入侵和本地木本生长和竞争的影响[D].武汉:华中农业大学,2018
[31] 胡可易,柳丽娜,李雪涛,等.经营干扰对毛竹林土壤肥力及胸径分布的影响[J].浙江林业科技,2013,33(5):52-56
[32] 洪宜聪. 毛竹纯林和混交林竹笋-幼竹高生长差异分析[J]. 西南林业大学学报, 2013, 33(5): 64-68
[33] 樊江文, 钟华平, 杜占池,等. 草地植物竞争的研究[J]. 草业学报, 2004(3):1-8
[34] Grime J P . Vegetation classification by reference to strategies[J]. Nature, 1974, 250:26-31
[35] 趙雨虹.毛竹扩张对常绿阔叶林主要生态功能影响[D].北京:中国林业科学研究院,2015
[36] 杨清培,杨光耀,宋庆妮,等.竹子扩张生态学研究:过程、后效与机制[J].植物生态学报,2015,39(1):110-124
[37] 徐晓,杨丹.湖南省马尾松林生物总量的空间分布与动态变化[J].中南林业科技大学学报,2012,32(11):73-78
[38] 杨清培,王兵,郭起荣,等.大岗山毛竹扩张对常绿阔叶林生态系统碳储特征的影响[J].江西农业大学学报,2011,33(3):529-536
[39] 杜莹,鲍永新,裘子炎,等.模拟氮沉降对毛竹非结构性碳的影响[J].林业科学,2017,53(7):10-17
[40] 刘宁. 氮沉降和生物炭添加对毛竹生理特性的影响[D]. 临安:浙江农林大学,2019
[41] Guo Y Y,Xing Y J,Wang J Y,et al.Sequestration of atmospheric CO2 in boreal forest carbon pools in northeastern China: Effects of nitrogen deposition[J]. Agricultural and Forest Meteorology,2018,248:70-81
[42] 陶晨悦,邵珊璐,史文辉,等.氮沉降对干旱胁迫下毛竹实生苗生物量和保护酶活性的影响[J].林业科学,2019,55(9):31-40
