不同UV-B辐射增幅对稻田土壤酶活性、活性有机碳含量及温室气体排放的影响
2021-09-09陈思王灿李想李明锐湛方栋李元祖艳群何永美
陈思,王灿,李想,李明锐,湛方栋,李元,祖艳群,何永美
云南农业大学资源与环境学院,云南 昆明 650201
紫外线(ultraviolet,UV)依据波长,分为3类:UV-A(315—400 nm)、UV-B(280—315 nm)和UV-C(100—280 nm),其中UV-B辐射危害最大(Sato et al.,1996)。氧分子通过相互作用形成臭氧集聚于平流层,能吸收全部UV-C和部分UV-B。由于氟氯碳化物(CFCs)等人造化学物质的大量使用,导致到达地表的生物有效UV-B辐射强度增加。自1989年《蒙特利尔议定书》签订实施以来,大气中的氟氯烃含量下降,2013—2016年南极平流层中无机氯年平均递减率约为0.8%(Strahan et al.,2018),从1998年起,南极臭氧层在逐渐恢复,但低纬度地区平流层臭氧总量在持续下降(Ball et al.,2018),所以到达地表的UV-B辐射水平仍不断增强。
全球稻田CH4排放量占全球农田总温室气体排放量的48%(Kimberly et al.,2017),中国作为世界最大的稻米生产国,稻田CH4排放量占全世界总排放量的 29.2%,对全球温室效应的贡献率约为20%(Kirschke et al.,2013;Chen et al.,2021)。稻田是重要的农田生态系统,也是全球碳循环中重要的碳库。稻田CH4的排放与土壤碳转化酶活性、活性有机碳含量密切相关。稻田土壤碳转化酶对土壤活性有机碳的含量有重要影响,推动土壤含碳物质的转化,且土壤碳转化酶活性和碳排放量反映土壤活性有机碳的有效性(Burns et al.,2013)。稻田土壤活性有机碳主要来源于凋落物、腐殖质的降解产物和根系分泌物等,其含量会随着水稻生育期和耕作方式的不同以及外界环境变化等因素发生变化(吴家梅等,2017)。而且土壤微生物群落随水稻还田秸秆的分解大量繁殖,使土壤蔗糖酶、多酚氧化酶和纤维素分解酶活性升高(Li et al.,2020),既减少碳的流失,也促进土壤有机碳的转化,显著提高土壤中活性有机碳的含量,为CH4的产生提供了丰富的底物,促进稻田CH4、CO2等温室气体的排放(Henning et al.,2021)。
UV-B辐射增强对陆地生态系统有显著影响,在凋落物分解和土壤物质循环过程中起着重要的调控作用(Erickson et al.,2015)。UV-B辐射增强影响植株根系生长和根系分泌物的产生,根际环境发生改变(Mahmood et al.,2013),导致土壤微生物数量增加,进而影响微生物群落的结构和功能(何永美等,2016),同时UV-B辐射增强引起光化学反应,促进植物残渣在在光化学降解作用下释放出大量溶解性有机物并影响微生物活动(Kögel-Knabner et al.,2010),使土壤多酚氧化酶、蔗糖酶和纤维素酶活性升高,增加土壤微生物量碳和易氧化有机碳的含量,最终导致稻田 CH4、CO2和N2O排放量增加(Lou et al.,2016;Li et al.,2020),增加稻田CH4和N2O排放的综合增温潜势(肇思迪等,2017),但也有减少稻田CH4排放的报道(Lou et al.,2012;王灿等,2018)。然而,在不同UV-B辐射增幅下,对处于水稻生长期稻田土壤碳转化酶活性、活性有机碳含量以及温室气体排放影响的研究不够系统,有待进一步加强。
元阳梯田位于云南省红河州元阳县,特殊的地理结构和气候条件,形成全年淹水型水稻种植农田系统,常年淹水状态使稻田长期处于厌氧环境,有利于温室气体产生(Hou et al.,2013)。元阳梯田位于中国西南的云南高原,UV-B辐射较强,且UV-B辐射增强显著影响元阳梯田水稻生长期稻田土壤碳转化酶活性和活性有机碳含量,使稻田 CH4排放通量增加(徐渭渭等,2015;Li et al.,2020)。因此,本试验以元阳梯田为研究地点,人工模拟 UV-B辐射增强,在不同UV-B辐射增幅条件下(0、2.5、5.0、7.5 kJ·m−2),测定水稻生长期稻田土壤碳转化酶活性、活性有机碳含量以及CH4、CO2和N2O排放通量,探究自然光照与UV-B辐射增幅处理间的差异,分析不同 UV-B辐射增幅对水稻生长期稻田土壤碳转化酶活性、活性有机碳含量以及温室气体排放的影响。
1 材料和方法
1.1 试验地介绍
元阳梯田随山势地形开垦,在平缓的坡面上形成梯田3000多级,面积近1.33×104 hm2,是哈尼族人经过1500多年开垦得到的自然-人工复合生态系统。该地处于亚热带季风气候,年均降水量1400 mm,年平均温度在16 ℃左右,全年日照时间长达1670 h。试验地位于元阳县元阳梯田的核心区新街镇箐口村,其平均海拔 1600 m,坡度 26°,坡向NE25°,该地 UV-B 辐射背景值 10 kJ·m−2·d−1(He et al.,2016)。土壤pH值5.32,有机质质量分数26.8 g·kg−1,全氮 1.76 g·kg−1,全磷 0.45 g·kg−1,全钾 16.20 g·kg−1,碱解氮 78.44 mg·kg−1,速效钾 16.52 mg·kg−1,速效磷 15.26 mg·kg−1。
1.2 试验设计与田间管理
试验材料选择元阳当地水稻品种白脚老粳,2017年3月播种育苗,5月将秧苗移栽至试验区内,共布置12个小区,每一个小区面积2.25 m×3.9 m,每一个试验区移植14行、16列水稻丛。在水稻生长期间进行稻田常规管理,不使用任何农药和化肥,栽培期间始终保持淹水状态,全年水深一般维持在10 cm左右。
设置 0、2.5、5.0、7.5 kJ·m−2UV-B辐射递增处理,自然光为对照处理,相当于0%、10%、20%、30%的臭氧衰减量,每个处理3个重复。从水稻秧苗移栽稻田成活后至水稻收获,每天10:00—17:00共照射7 h(阴雨天除外)。试验小区中间每行水稻上方悬挂1支40 W UV-B灯管(北京,UV308,光谱为280—320 nm),共悬挂10支,灯管高度距水稻顶端60、40、20 cm,分别为UV-B辐射增强2.5、5.0、7.5 kJ·m−2,自然光照处理仅悬挂灯架,随水稻生长改变灯管距水稻顶端的高度调节辐照强度。
1.3 样品采集及测定方法
在水稻分蘖期、拔节期、孕穗期、抽穗期、成熟期采集水稻周围土壤,使用土样采集器用“S”布点法采集稻田表层0—20 cm的土壤,每小区采集1 kg新鲜土样,多余土壤按照四分法去掉,装于密封袋中做好标记,土样放入低温储存箱并迅速带回试验室分析(赵炳梓等,2011;耿玉清等,2012)。
土壤纤维素酶活性:称取过0.25 mm筛风干土样,加甲苯、醋酸盐缓冲液和羧甲基纤维素溶液,37 ℃培养72 h,过滤定容后加50 mL蔥酮试剂后进行比色(关松荫,1986)。
土壤β-葡萄糖苷酶活性:称取过0.25 mm筛风干土样,加甲苯、MUB(pH 6.0)和葡葡糖苷溶液,37 ℃培养1 h后加CaCl2和THAM,于400 nm处比色(关松荫,1986)。
土壤多酚氧化酶活性:称取过0.25 mm筛风干土样,加邻苯三酚溶液,30 ℃培养2 h后加柠檬酸-磷酸缓冲液和35 mL乙醚萃取后比色(关松荫,1986)。
土壤蔗糖酶活性:称取过0.25 mm筛风干土样,加蔗糖、磷酸缓冲液和甲苯,37 ℃培养24 h后过滤,滤液加3, 5-二硝基水杨酸,沸水浴加热5 min,于508 nm处比色。每一土样需做无基质对照(关松荫,1986)。
溶解性有机碳(Dissolved organic carbon,DOC)含量:称取新鲜土样,加硫酸钾溶液浸提,滤液加重铬酸钾-硫酸混合液,邻啡罗啉作指示剂,根据硫酸亚铁标准液滴定量,计算得土壤 DOC含量(盛浩等,2015)。
易氧化有机碳(Labile organic carbon,LOC)含量:称取新鲜土样,加高锰酸钾氧化,离心取上清液,用分光光度法于565 nm比色,根据高锰酸钾的消耗量,计算LOC含量(Blair et al.,1995)。
微生物量碳(Microbial biomass carbon,MBC)含量:称取土样氯仿熏蒸24 h,加硫酸钾溶液浸提,加重铬酸钾-硫酸混合液,邻啡罗啉作指示剂,根据硫酸亚铁标准液滴定量,计算得土壤MBC含量。需设置空白对照(李振高等,2008)。
气体排放通量的测定:参照娄运生等(2012)气体采集方法,使用静态箱收集。采集气体时间确定在09:00—11:00之间,进行气体收集时先将采气箱置于水稻上,利用稻田自身的淹水层进行密封以保证采集箱的密封性,盖上采气箱盖子后立即向500 mL的真空铝箔气体袋中泵入采气箱中的气体,采气时间在采气箱罩在水稻上方的 0、10、20、30 min,每10分钟进行一次采样。
使用Agilent 7890B气相色谱仪来测量CH4和CO2的排放通量。检测器FID参数为:加热器温度为 210 ℃,H2流量为 40 mL·min−1,空气流量为 400 mL·min−1,实验柱的温度设置 50 ℃;色谱柱流量设置为 2.500 mL·min−1。
计算公式:
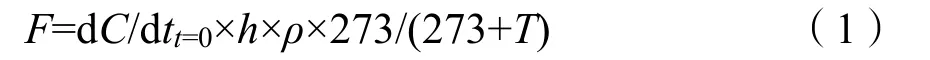
式中:
F为气体排放通量;
h为采样箱高度;
T为箱内温度;
ρ为标准状况下气体的密度;
dC/dtt=0为单位时间内气体浓度线性变化率。
全球增温潜能(global warming potential,GWP)是用来评价各种温室气体对气候变化影响相对能力的一个参数。以百年时间单位计,将CO2气体的GWP值设置为1,CH4和N2O气体的GWP分别是CO2的 25 倍和 298 倍(Kou et al.,2015;Chen et al.,2021)。因此,将总温室气体排放量(g·m−2)的变暖潜能转换为 CO2当量,稻田 CH4、N2O和 CO2排放的全球增温潜能计算方法为:
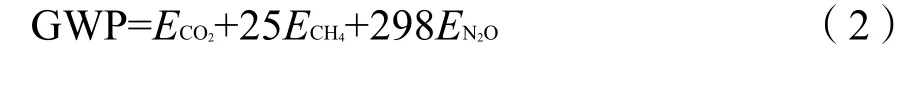
1.4 数据处理
采用Excel软件进行数据处理,SPSS 21.0进行差异性和相关性分析,OriginPro 9.0进行绘图。
2 结果与分析
2.1 不同 UV-B辐射增幅对水稻生长期稻田土壤碳转化酶活性的影响
4种有机碳转化酶活性随生育期的变化而发生变化,但不同酶之间存在显著差异(见图1)。与自然光相比,2.5 kJ·m−2UV-B辐射处理导致孕穗期和抽穗期纤维素酶活性显著升高;5.0 kJ·m−2UV-B辐射处理导致分蘖期、拔节期、孕穗期纤维素酶活性显著升高,升高幅度为 15.4%—37.7%;7.5 kJ·m−2UV-B辐射处理使分蘖期、抽穗期和成熟期纤维素酶活性显著降低,降幅为21.7%—24.2%;且该处理下成熟期纤维素酶活性显著低于其他时期。

图1 不同UV-B辐射增幅对稻田土壤碳转化酶活性的影响Fig. 1 Effects of different UV-B radiation levels on the activity of soil carbon invertase in the rice paddy
UV-B辐射增幅增加导致分蘖期、拔节期和抽穗期 β-葡萄糖苷酶活性显著降低,且与自然光相比,5.0 kJ·m−2UV-B辐射处理导致β-葡萄糖苷酶活性显著降低,降幅为 10.7%—23.3%;7.5 kJ·m−2UV-B辐射处理导致拔节期、抽穗期β-葡萄糖苷酶活性显著降低28.7%、44.5%,且该处理下抽穗期β-葡萄糖苷酶活性显著低于其他时期。
总体来看,土壤多酚氧化酶活性随UV-B辐射增幅逐渐增加而降低,且与自然光相比,7.5 kJ·m−2UV-B辐射处理导致水稻生长期多酚氧化酶活性显著降低,最大降幅为 29.4%;自然光和 2.5 kJ·m−2UV-B辐射处理下,分蘖期和拔节期土壤多酚氧化酶活性显著高于其他3个时期。
随UV-B辐射增幅增加,水稻生长后期(孕穗期、抽穗期、成熟期)土壤蔗糖酶活性显著降低,且7.5 kJ·m−2UV-B辐射处理下抑制作用最显著。5.0、7.5 kJ·m−2UV-B辐射处理导致孕穗期土壤蔗糖酶活性显著低于其他生长期。由双因素分析可知,水稻生长期和UV-B辐射对土壤4种有机碳转化酶活性均有显著影响,且存在交互作用。
2.2 不同 UV-B辐射增幅对水稻生长期稻田土壤活性有机碳含量的影响
由图2可知,不同UV-B辐射增幅稻田土壤活性有机碳含量随着生长期的不同呈现出不同的变化趋势。与自然光相比,5.0 kJ·m−2UV-B辐射处理导致分蘖期、抽穗期和成熟期土壤 DOC含量显著增加,增幅范围为17.5%—47.9%。自然光、2.5和7.5 kJ·m−2UV-B辐射处理下,成熟期土壤DOC含量显著低于其他时期。

图2 不同UV-B辐射增幅对稻田土壤活性有机碳含量的影响Fig. 2 Effects of different UV-B radiation levels on the content of active organic carbon in the rice paddy
随UV-B辐射增幅增加,水稻拔节期、孕穗期和抽穗期土壤LOC含量显著减少,但UV-B辐射增幅处理间无显著差异;与自然光相比,7.5 kJ·m−2UV-B辐射处理导致成熟期土壤LOC含量显著减少21.1%。自然光和2.5 kJ·m−2UV-B辐射处理下,孕穗期土壤LOC含量显著高于其他生长期。
当UV-B辐射增幅增加,稻田土壤MBC含量显著减少。与自然光相比,7.5 kJ·m−2UV-B辐射处理下水稻分蘖期、拔节期、孕穗期和成熟期土壤MBC含量显著减少,分别减少 18.2%、24.5%、22.6%、35.9%;自然光条件下,孕穗期和成熟期土壤MBC含量显著高于其他时期;分蘖期土壤MBC含量显著低于其他时期。双因素分析发现,水稻生长期和UV-B辐射对土壤DOC、MBC含量均有显著影响,但没有交互作用。
2.3 不同 UV-B辐射增幅对水稻生长期稻田温室气排放通量的影响
由图3可知,水稻分蘖期、拔节期、孕穗期和抽穗期,5.0、7.5 kJ·m−2UV-B辐射处理导致CH4排放通量显著减少,最大降幅为30.6%;自然光和3个UV-B辐射增幅处理,抽穗期CH4排放通量显著高于其他时期。综合水稻整个生育期,CH4排放通量的变化趋势基本一致,其排放通量随UV-B辐射增幅增加而降低。

图3 不同UV-B辐射增幅对稻田温室气排放通量的影响Fig. 3 Effects of different UV-B radiation levels on the greenhouse gas emission fluxes in the rice paddy
分蘖期,7.5 kJ·m−2UV-B 辐射处理导致 CO2排放通量显著减少,而UV-B辐射增幅处理间无显著差异;与自然光照相比,5.0 kJ·m−2UV-B辐射处理导致水稻拔节期、孕穗期和抽穗期CO2排放通量显著增加,分别增加 25.0%、17.7%、30.7%。5.0 kJ·m−2UV-B辐射处理下,抽穗期CO2排放通量显著高于其他时期。
与自然光相比,7.5 kJ·m−2UV-B辐射处理导致拔节期、孕穗期和成熟期N2O排放通量显著减少,5.0 kJ·m−2UV-B辐射处理使拔节期和成熟期 N2O排放通量显著增加。2.5、5.0 kJ·m−2UV-B辐射处理导致孕穗期 N2O排放通量显著高于其他时期,7.5 kJ·m−2UV-B辐射处理导致拔节期N2O排放通量显著低于其他时期。双因素分析表明,水稻生长期和UV-B辐射对稻田CH4、CO2、N2O排放通量均存在极显著影响,且存在交互作用。
2.4 不同 UV-B辐射增幅对稻田温室气体增温潜能的影响
不同UV-B辐射增幅对水稻生长期稻田土壤3种温室气体GWP的影响如图4所示。与自然光相比,7.5 kJ·m−2UV-B辐射处理导致水稻生长期稻田GWP显著降低,降幅范围为 10.3%—16.2%。5.0 kJ·m−2UV-B辐射处理导致孕穗期稻田GWP显著高于其他时期。双因素分析表明,水稻生长期和UV-B辐射对稻田GWP均有显著影响,但没有交互作用。
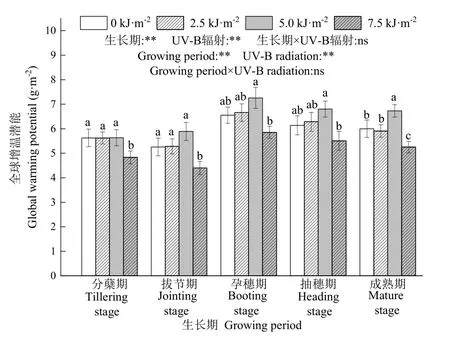
图4 不同UV-B辐射增幅对稻田3种温室气体增温潜能的影响Fig. 4 Effects of different UV-B radiation levels on the global warming potential of three greenhouse gases in the rice paddy
2.5 相关性分析
由相关性分析可得(表1),经UV-B辐射增强处理,多酚氧化酶与 MBC和 LOC呈显著正相关(P<0.05),与DOC呈显著负相关(P<0.05)。

表1 水稻生育期稻田土壤碳转化酶活性与活性有机碳含量的相关性Table 1 Correlation coefficients between carbon invertase activities and active organic carbon contents in the rice paddy
由表 2可知,MBC与 CH4呈极显著正相关(P<0.01),LOC与 MBC呈显著正相关,DOC与CH4呈显著负相关(P<0.05)。

表2 水稻生育期土壤活性有机碳含量与碳排放通量的相关性Table 2 Correlation coefficients between soil active organic carbon contents with carbon gas emissions in the rice paddy
3 讨论
3.1 UV-B辐射增强对土壤碳转化酶、活性有机碳转化的影响与机理
土壤酶是土壤功能和生物活性的重要指标,在土壤有机质的形成与降解、养分循环和外源生物的分解过程中起着重要作用(李增强,2018)。其中,土壤蔗糖酶催化蔗糖水解为葡萄糖和果糖(Gu et al.,2009),纤维素酶和 β-葡萄糖苷酶能够将难溶的纤维素类物质分解为易溶于水的糖类物质(Wickings et al.,2012),多酚氧化酶主要参与土壤中木质素等复杂物质的氧化降解(Sinsabaugh,2010),这些碳转化酶有助于促进土壤含碳物质转化,促进活性有机碳的生成。
土壤有机碳库可作为土壤管理短期变化的敏感指标,土壤MBC、DOC和LOC是土壤有机碳库中的重要组成部分(Song et al.,2012)。土壤MBC代表了土壤中活性微生物生物量的大小,很大程度上取决于底物碳的供应(Burns et al.,2013),DOC和 LOC是由土壤微生物驱动下的植物残渣和有机物分解以及根系分泌产生的,它们都与土壤有机碳的积累和降解过程密切相关(Bernd et al.,2003;Wang et al.,2013;Yang et al.,2013)。而且,土壤碳转化酶活性会随着活性有机碳组分含量的变化而出现相同变化(李增强等,2018)。
本试验发现随UV-B辐射增强,土壤MBC和LOC含量减少,而且多酚氧化酶和蔗糖酶活性的变化与MBC和LOC含量变化基本一致。多酚氧化酶活性与MBC和LOC含量呈显著正相关,与DOC含量呈显著负相关,这与前人研究结果相一致(王灿等,2018)。同时试验还发现 UV-B辐射增强使土壤DOC含量增加,因为UV-B辐射增强使水稻根际微生物数量和根系分泌物量增加,促进植物残体、秸秆分解成有机质,并在转化过程中释放大量DOC(He et al.,2016;Li et al.,2020)。
臭氧衰减导致的地表 UV-B辐射增强,是影响稻田生态系统的重要环境因子之一(Wargent et al.,2013)。紫外线不能穿透5 mm以下的土壤,对微生物群落的影响是间接的,主要是通过直接影响植物生长和生理代谢,损伤水稻叶片,降低叶片光合能力,减少水稻植株生物量,使得水稻对有效态碳、氮的吸收减少,从而影响微生物群落(Bao et al.,2018;李想等,2018)。在水稻生长期,UV-B辐射增强会改变水稻根系分泌物的种类和分泌量,导致根际微生物的群落组成与数量发生变化(何永美等,2016),间接影响根际土壤碳转化酶活性与活性有机碳含量。同时,有研究发现 UV-B辐射增强对植株根系分泌物及土壤微生物的影响并不都是促进或抑制,而且植物对 UV-B辐射的响应存在低促高抑的现象,这可能与辐射强度和环境因素等试验条件差异有关(李元等,1999;Robson et al.,2004;Lou et al.,2016;何永美等,2016;Gao et al.,2019)。
3.2 UV-B辐射增强对稻田温室气体排放的影响与机理
土壤碳转化酶活性、土壤碳库中 DOC、LOC和 MBC均与CH4的排放密切相关(Zhan et al.,2011),而且产甲烷菌主要以LOC为底物产生CH4(吴家梅等,2013)。稻田土壤CH4主要通过水稻植株的通气组织向大气排放(Bhattacharyya et al.,2019),UV-B辐射增强可显著降低大多数湿地植物的光合作用、叶片横截面和通气组织气孔比例(Niemi et al.,2010;Robson et al.,2015;Wagh et al.,2019),是导致稻田 CH4排放量减少的一个原因。另外,本研究中CH4排放通量与MBC含量呈显著正相关性,可能是随着UV-B辐射增强根际微生物活性和根系分泌物量减少,导致土壤碳转化酶活性与活性有机碳含量降低,从而减少稻田CH4的排放量(Lou et al.,2012;王灿等,2018)。但这方面的研究结果存在差异(Lou et al.,2016;胡正华等,2011;徐渭渭等,2015),这种差异与试验地和所用水稻品种有关,对UV-B辐射敏感的作物种间和种内都存在很大的差异性(何永美等,2012)。
土壤呼吸强度与土壤微生物数量、种类和活性关系密切,微生物的理化作用以及土壤中有机质含量等因素都能影响CO2排放量(李海防等;2007;Hu et al.,2010)。UV-B辐射增强促进凋落物分解和土壤呼吸作用,增加CO2排放通量(柳淑蓉等,2012)。而且,气孔控制着叶片与大气之间 CO2和水汽的扩散、传导,UV-B辐射增强会导致植物气孔导度减小,增加气孔对外界环境特别是空气湿度的敏感性,减少植株对CO2的吸收,UV-B辐射增加促进凋落物分解矿化生成 CO2,最终导致稻田土壤 CO2排放量增加(郑有飞等,2011;褚润等,2017)。同时,稻田土壤N2O的产生主要取决于硝化细菌和反硝化细菌对无机氮、根残留物和渗出液的硝化、反硝化作用(Kou et al.,2015),UV-B辐射通过影响土壤中有效 N、土壤微生物碳氮量、土壤微生物C/N,使土壤硝酸还原酶、蛋白酶活性升高,促进了NH4+-N向NO3−-N的转化,从而促进了N2O的排放(蒋静艳等,2006;肇思迪等,2017;张彦雪等,2020)。由于元阳梯田特殊的种植模式和气候条件,长期的淹水厌氧状态以及降雨量的增减都会影响水稻温室气体的排放量。温室气体排放对土壤碳氮库和GWP值具有重要影响(肇思迪等,2017),UV-B辐射增强抑制了稻田土壤碳转化过程,使3种温室气体的排放量减少,进而导致稻田3种温室气体GWP值降低。
4 结论
随着UV-B辐射增幅增加,水稻生长期稻田土壤β-葡萄糖苷酶、多酚氧化酶和蔗糖酶活性降低,导致稻田土壤MBC、LOC含量减少,抑制稻田土壤CH4排放,但促进了CO2、N2O排放;从CH4、CO2和N2O综合增温潜能看,高强度UV-B辐射能降低3种温室气体GWP。
