黔西北黄杉群落物种多样性的海拔梯度格局
2021-09-09何斌李青陈群利李望军游萍
何斌,李青,陈群利,李望军,游萍
贵州省普通高等学校生物资源开发与生态修复特色重点实验室/贵州工程应用技术学院生态工程学院,贵州 毕节 551700
物种多样性是研究植物群落结构与功能的物质基础(Curtis et al.,1951;张晓龙等,2017),不仅能够反映植物群落对生态环境的响应,而且可以有效评价生态系统的稳定性(刘维暐等,2012)和健康状况(Rad et al.,2009)。物种多样性的空间分布格局及其潜在机制一直是生态学和植物地理学研究的热点问题之一(Whittaker,1960;Macarthur,1972),受多个生态梯度(如经纬度、海拔、研究尺度等)的影响(Palmer,1992;黄建辉,1994),其中海拔梯度被认为是一个关键性的影响因子(Debano et al.,1990;Zimmerman et al,1999)。因此,在全球气候变暖的背景下,加强物种多样性的海拔梯度格局研究,对于揭示物种多样性的环境梯度变化规律具有重要意义(郝建锋等,2014)。
植物群落物种多样性的海拔梯度格局常常与研究区的地理位置、环境条件、研究对象和研究尺度等有关(Ghimire et al.,2006;方精云等,2004;吴昊等,2012)。已有研究表明,不同地区山地物种多样性的海拔梯度格局不同,热带地区以单调下降趋势为主,亚热带地区以先上升再下降趋势为主,温带地区变化趋势不确定(Ohsawa,1991;Ohsawa,1995;Liu et al.,2007;方精云,2004;沈泽昊等,2004;王志恒等,2004)。低海拔地区降水和人为干扰是限制物种多样性的关键因子(Wang et al.,2007),高海拔地区则为温度(Whittaker et al.,1975)。Beck et al.(2008)认为山地水热条件的综合效应是造成物种多样性单峰格局的主要原因,而Begon et al.(1986)认为温度是造成物种多样性随海拔梯度单调下降的主要原因;Lomolino(2001)和Rahbek(2005)认为物种多样性的海拔梯度格局存在不确定性,取样时的最低海拔梯度和不同海拔梯度的地形变化均可导致物种多样性单调下降的海拔梯度格局现象;Wu et al.(2014)研究认为进化因素也是造成物种多样性海拔格局的因素之一。由此可见,目前有关山地物种多样性的海拔梯度格局以及机制还没有形成统一的定论。因此,开展不同植物类群、不同山体和不同区域物种多样性海拔格局的案例研究,对阐明物种多样性与海拔之间的关系具有重要意义,有利于更好地理解山地植物对环境变化的适应(Kitayama,1992;Grytnes et al.,2002)。
随着全球经济活动的加剧,人类对植物资源的过渡利用和生态环境的破坏,导致大量物种濒于灭绝,特别是近几十年来,许多濒危物种的分布区迅速缩小。在这种背景下,濒危物种多样性的研究得到了众多生态学家的关注,并成为重点研究对象。黄杉(Pseudotsuga sinensis)属松科(Pinaceae)黄杉属(Pseudotsuga),为第三纪孑遗植物,中国特有种,被列为国家Ⅱ级重点保护植物,在全国分布范围狭窄,仅在贵州、滇东北、滇中、川东、川西南、湘西、鄂西等地呈零星分布(熊斌梅等,2017)。在贵州省威宁县分布的黄杉纯林,面积约373.2 hm2,是国内罕见的保存比较完整、面积较大的黄杉群落。由于黄杉林是黔西北地区喀斯特山地森林生态系统的一个重要组成部分,研究黄杉群落物种多样的垂直分布格局,不仅对探讨其濒危机制及生物多样性保护具有重要的意义,而且有利于揭示亚热带喀斯特山地森林生态系统的结构、演替和功能。目前,有关黄杉的研究较少,且主要集中在群落结构(左家哺,1995;熊斌梅等,2017)、种群动态(孟广涛等,2008)和生态评价(胡文彬,2015)等方面。基于此,本文以黄杉群落为研究对象,通过典型群落样地调查,分析群落物种组成、植物区系成分、物种多样性对海拔变化的响应,旨在探讨,(1)植物区系成分和物种多样性的海拔梯度格局?(2)物种α多样性各指数间是否存在相关性?(3)灌木层和草本层的植物多样性格局是否一致?以期为喀斯特山地生物多样性保护和濒危植物的保育提供参考,也为研究喀斯特山地森林植物群落的稳定性机制奠定基础。
1 研究区概况与方法
1.1 研究区概况
研究区位于贵州省威宁县西南部的哲觉、黑石、麻乍等6个乡镇,属中亚热带季风气候,年均温10.5—3.5 ℃,年均降雨量1000 mm左右,年总日照时数1530—1926 h。地形复杂、地势起伏大,坡度10°—30°之间,土壤母质以紫色砂岩和砂页岩为主,主要为黄壤、黄棕壤和紫色土,呈酸性(pH 5.5—6.0)。研究区内植被类型主要有黄杉林、云南松(Pinus yunnanensis)林、华山松(P. armandii)林、金丝桃(Hypericum monogynum)灌丛、马桑(Coriaria nepalensis)灌丛、西南栒子(Cotoneaster franchetii)灌丛、滇榛(Corylus yunnanensis)灌丛、荚蒾(Viburnum dilatatum)灌丛和荒山草坡。
研究区内黄杉林分在20世纪50年代到80年代期间被当地群众大肆砍伐,林地面积减少了88.95%、蓄积减少了89.71%(黄鹤先,1989)。现有林分中,黄杉以中、幼林分居多,根据野外调查,主要有以下4种类型:黄杉纯林;黄杉、云南松混交林;黄杉、华山松混交林和黄杉散生木,且大部分为天然更新。
1.2 样地设置与调查
在对黄杉分布区全面踏查的基础上,结合实际情况,采用典型样地法于2019年7—10月从海拔1800 m的金斗乡到2290 m的哲觉镇,分别在6个海拔梯度上选择有代表性的黄杉林设置20 m×30 m的样地,每个海拔梯度2个重复,共12块样地,对乔木层植物进行每木检尺,同时记录样地的海拔、经纬度、坡度、地形等基本信息(表1)。在每个样地内,按梅花形选取5个5 m×5 m的灌木层样方和1 m×1 m的草本层样方,分别调查灌木植物的种类、株数(丛数)、高度和冠幅和草本植物的种类、株数(丛数)、平均高度和盖度。
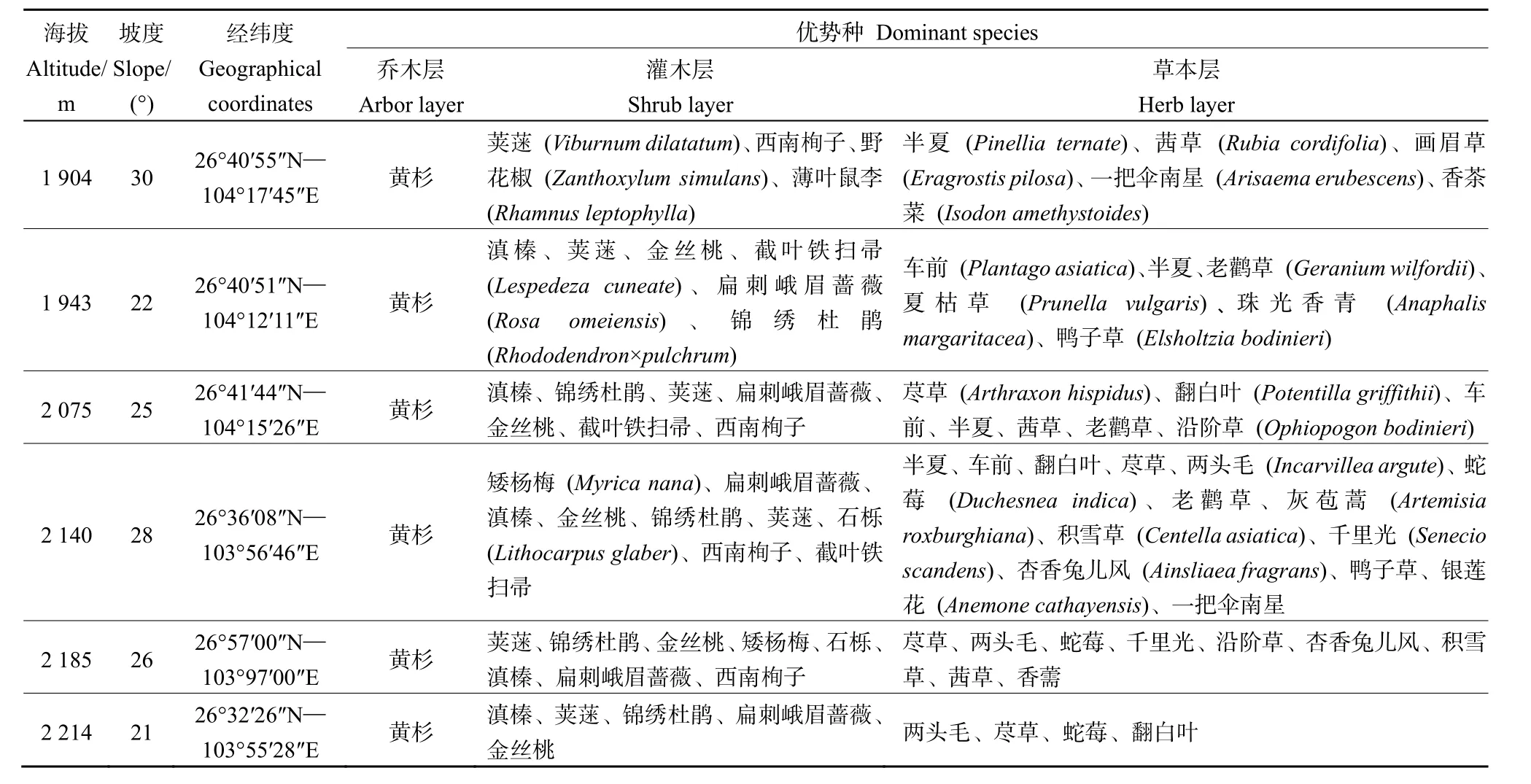
表1 研究区内黄杉林样地概况Table 1 General characteristics of the sample plot of P. sinensis forest in the study area
1.3 植物区系分析
参照《世界种子植物科的分布区类型系统》(吴征镒等,2003)和《中国种子植物属的分布区类型》(吴征镒,1991)对中国种子植物分布区类型的划分,进行统计分析。
1.4 物种多样性分析
参考马克平等(1995)和方精云等(2004)的生物多样性测度方法,本文从α多样性和β多样性两个方面分析海拔梯度对物种多样性的影响。
灌木层和草本层重要值(IV)计算公式为:

式中:
Hr为相对高度;
Cr为相对盖度;
Fr为相对频度。
(1)α多样性指数:
物种丰富度指数(S):

式中:
N为样地内出现的物种数。
Shannon-Wiener多样性指数:

Simpson优势度指数:

Pielou均匀度指数:

式中:
Pi为物种i的重要值。
(2)β多样性指数:
Sørenson物种相似性指数(Cs):

Cody物种替代指数(βc):

式中:
a、b分别为任意两个海拔梯度的物种数;
c为两海拔梯度间的共有物种数。
1.5 数据分析
运用SPSS 19.0统计软件进行以下分析:(1)单因素方差分析(One-Way ANOVA),以海拔梯度为自变量,以不同层次的物种多样性指数为因变量进行方差分析和显著性检验;(2)黄杉群落不同层次物种多样性的 Pearson相关性分析。应用Microsoft Excel 2016进行绘图。
2 结果与分析
2.1 植物群落的物种组成
本次调查,样地中共记录种子植物50科95属124种。由表2可知,含有10个以上物种的科有2科,占总物种数比例为 23.39%,分别为蔷薇科(Rosaceae)16种和菊科(Compositae)13种;含5—9个物种的科共有 5科,占总物种数的比例为25.81%,分别为杜鹃花科(Ericaceae)8种、禾本科(Gramineae)7种、壳斗科(Fagaceae)6种、豆科(Leguminosae)6种和唇形科(Labiatae)5种。上述7个科仅占总科数的14%,但是所包括的物种占总物种数的49.2%,说明这7个科的植物在黄杉群落区系组成中起主导作用。含4个物种及以下的科有43科,其中31科只含有1个物种,占总物种数的50.8%,但是占总科数的86%。
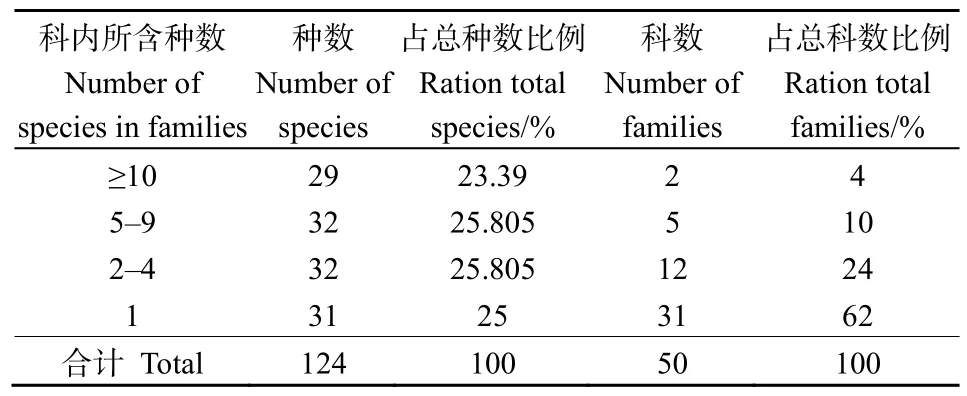
表2 黄杉群落种子植物科内种的组成Table 2 Composition of species in families of seed plants in the P. sinensis community
由表3可知,群落内各科所含属数差异也很明显。含有10属以上的科只有1科,为菊科,占总科数的2%,占总属数的11.58%;含有4—9属的科共有5科,占总科数的10%,占总属数的30.53%,分别为蔷薇科9属、禾本科6属、豆科5属、唇形科5属和桦木科4属;含3属及以下的科共有44科,占总科树的88%,占总属数的57.89%,其中33科只有1属。
基于语文核心素养要求的作文自改教学是一种行之有效的教学方法,对教学策略的探索也是有益的,但本文中的策略设计还不够完美,对实际的教学问题的考虑也还不够周全,希望能够在日后的探索中精益求精,尽善尽美。我相信,只要方向是正确的,即便此时仍旧行走在羊肠小道,只要坚持开拓,前方终会迎来坦途。
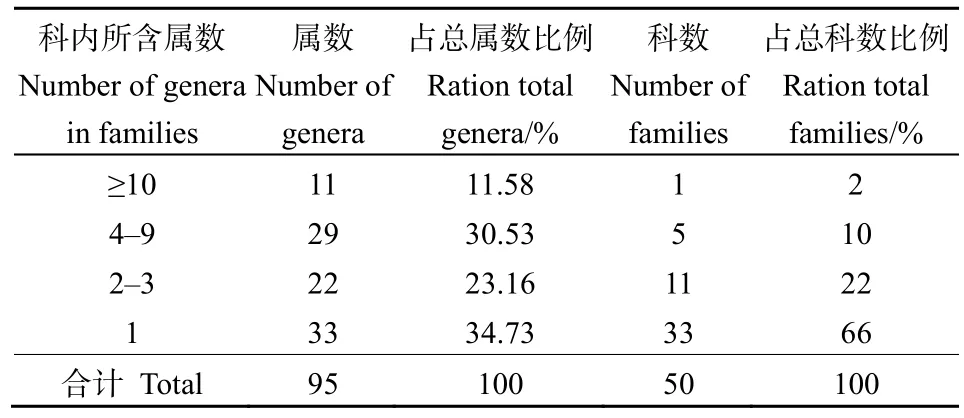
表3 黄杉群落种子植物科内属的组成Table 3 Composition of genera in families of seed plants in the P. sinensis community
2.2 植物区系成分
由表4可知,黄杉群落内种子植物科的地理成分较少,仅有6种分布区类型,其中世界分布最多,有19科,占总科数的38%,有禾本科、蔷薇科、菊科和唇形科等。热带地理成分共有20科,占总科数的40%,包括泛热带分布16科、东亚(热带、亚热带)及热带南美间断分布2科、热带亚洲至热带非洲分布1科和热带亚洲(印度—马来西亚)分布1科。温带地理成分中仅有北温带分布,共有11科,占总科数的22%。
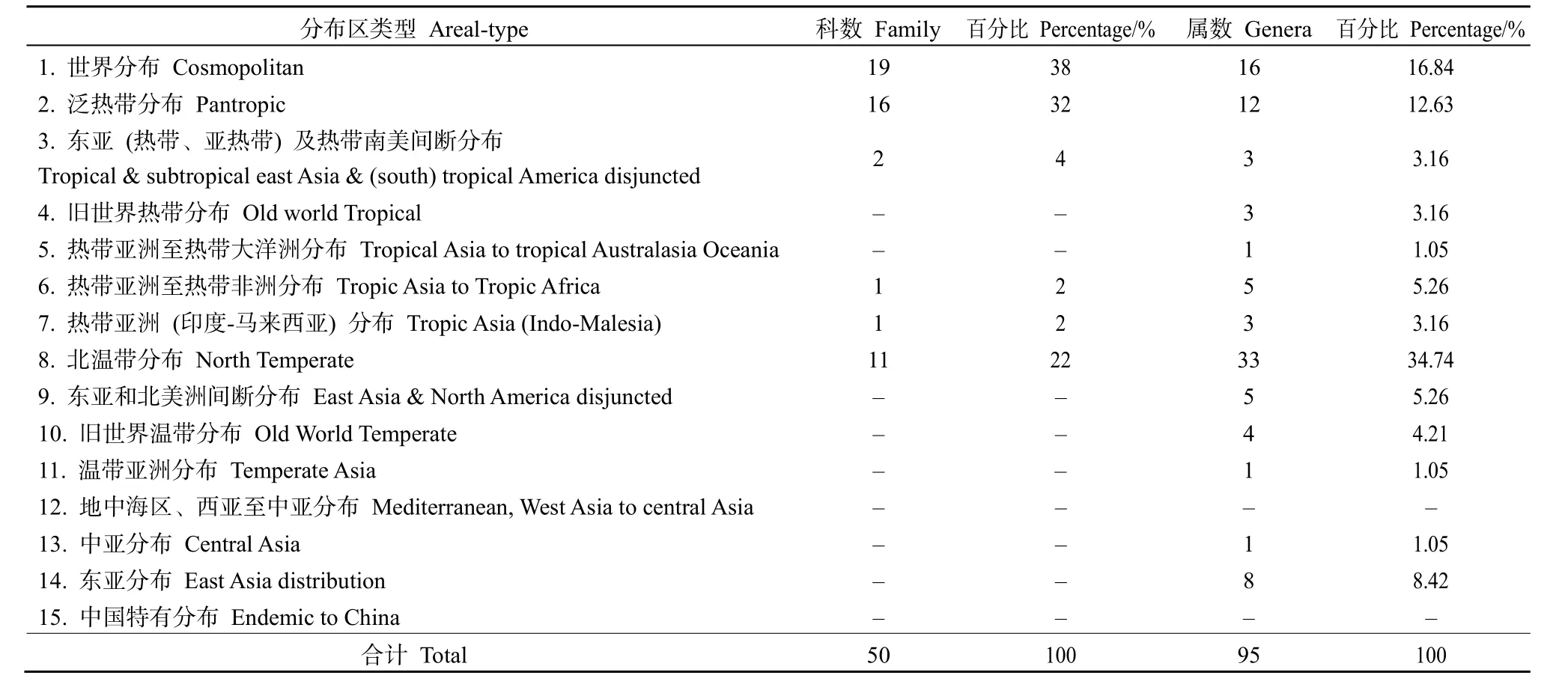
表4 种子植物科、属的分布区类型Table 4 The distribution type of families and genera in the flora of seed plants
在 15种分布区类型中,属的地理成分仅缺少了地中海区、西亚至中亚分布和中国特有分布,表明黄杉群落种子植物属的分布区类型复杂、多样。其中,北温带分布、世界分布和泛热带分布占据了群落的主体。世界分布型共有 16属,占总属数的16.84%,有金丝桃属(Hypericum)、悬钩子属(Rubus)、车前属(Plantago)和飞蓬属(Erigeron)等。热带地理成分共有27属,占总属数的28.42%。其中,泛热带分布(12属)占优势,占热带成分的44.44%,有醉鱼草属(Buddleja)、花椒属(Zanthoxylum)和薯蓣属(Dioscorea)等;热带亚洲至热带非洲分布有5属,占热带成分的18.52%,有铁仔属(Myrsine)和香茶菜属(Rabdosia)等;东亚(热带、亚热带)及热带南美间断分布有3属,占热带成分的 11.11%,有柃属(Eurya)和葎草属(Humulus)等;旧世界热带分布有3属,占热带成分的11.11%,有乌蔹莓属(Cayratia)和千金藤属(Stephania)等;热带亚洲(印度-马来西亚)分布有3属,占热带成分的11.11%,有润楠属(Machilus)和木荷属(Schima)等;热带亚洲至热带大洋洲分布有 1属,占热带成分的 3.71%,有栝楼属(Trichosanthes)。温带地理成分多于热带地理成分,共有52属,占总属数的54.74%。北温带分布占优势,有 33属,占总属数的 34.74%,有荚蒾属(Viburnum)、马桑属(Coriaria)、小檗属(Berberis)和栒子属(Cotoneaster)等;东亚分布有8属,占总属数的 8.42%,有栎属(Quercus)和半夏属(Pinellia)等;东亚和北美洲间断分布有 5属,占总属数的5.26%,有黄杉属(Pseudotsuga)和胡枝子属(Lespedeza)等;旧世界温带分布有4属,占总属数的4.21%,有火棘属(Pyracantha)和香薷属(Elsholtzia)等;温带亚洲分布有1属,占总属数的1.05%,有杭子梢属(Campylotropis);中亚分布有1属,占总属数的1.05%,有角蒿属(Incarvillea)。
2.3 物种多样性的海拔梯度格局
由于乔木层基本上为黄杉,偶尔有零星华山松(P. armandii)、云南松(P. yunnanensis)、川滇桤木(Alnus ferdinandi-coburgii)和栓皮栎(Quercus variabilis)分布,故下面主要分析灌木层和草本层物种多样性的海拔梯度格局。
2.3.1 α多样性的海拔梯度格局
从黄杉群落灌木层和草本层物种多样性的海拔梯度变化趋势图(图 1)中可以看出,随着海拔的升高,灌木层和草本层的物种丰富度指数 S、Shannon-Wiener多样性指数H、Simpson优势度指数D和Pielou均匀度指数J均呈单峰格局。其中,物种丰富度指数S、Shannon-Wiener多样性指数H和Simpson优势度指数D变动幅度较大,均在海拔2140 m处达到最大值;Pielou均匀度指数J变化较平缓,灌木层在海拔1943 m处达到最大值,而草本层在海拔2075 m处达到最大值。

图1 灌木层和草本层物种多样性的海拔梯度变化格局Fig. 1 Altitudinal gradient pattern of species diversity in shrub layer and herb layer
从图2中可以看出,不同海拔间,灌木层的物种丰富度、Shannon-Wiener多样性指数H、Simpson优势度指数D差异显著(P<0.05),均在海拔2140 m处出现最大值和海拔 1904 m处出现最小值;Pielou均匀度指数J差异不显著(P<0.05),最大值出现在海拔1943 m处,最小值出现在海拔2214 m处。不同海拔梯度间,草本层的物种丰富度、Shannon-Wiener多样性指数H、Simpson优势度指数D差异显著(P<0.05),均在海拔2140 m处出现最大值和海拔2214 m处出现最小值;Pielou均匀度指数J差异也显著(P<0.05),最大值出现在海拔2075 m处,最小值出现在海拔1904 m处。
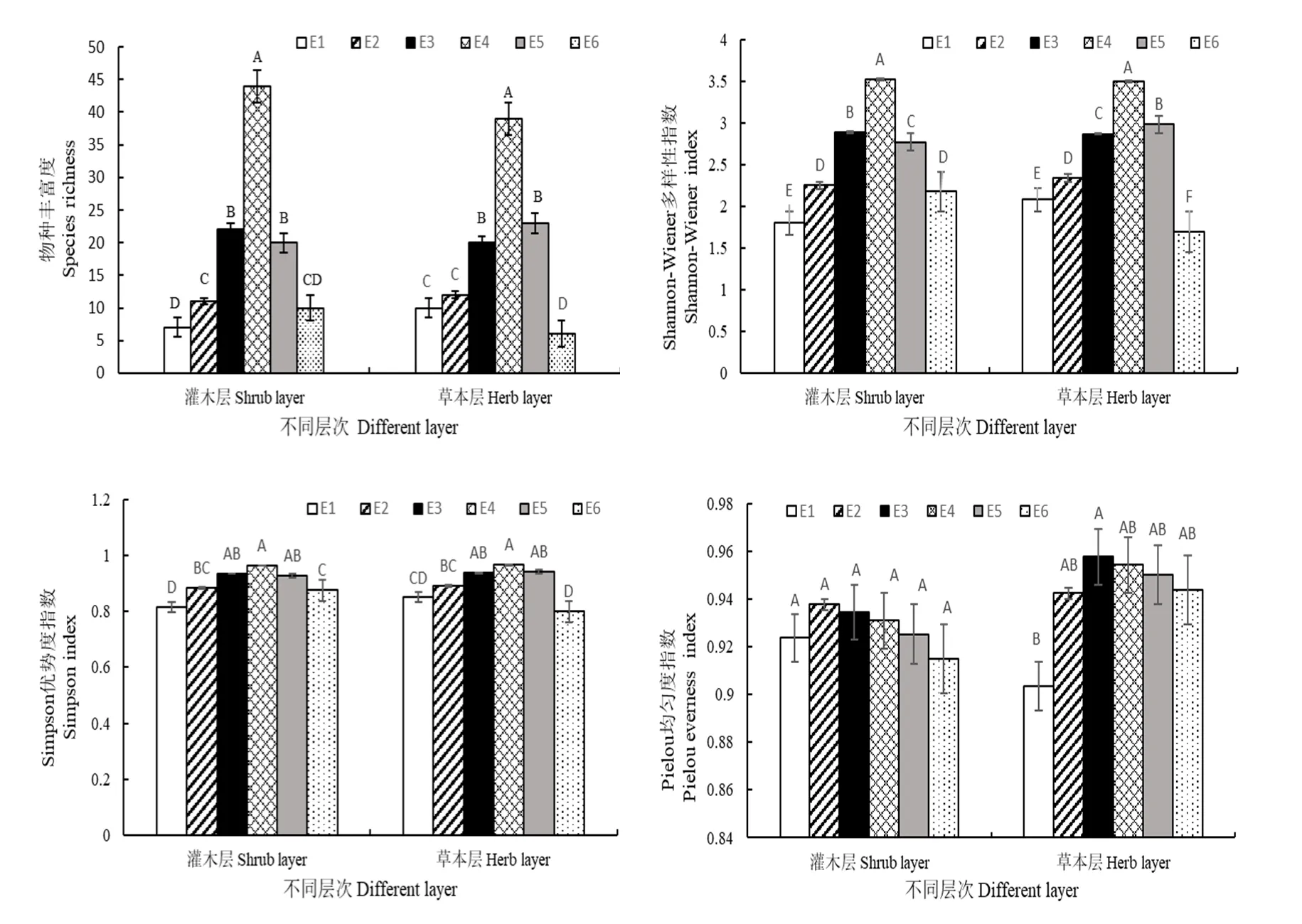
图2 不同海拔灌木层和草本层物种多样性的比较Fig. 2 Comparison of species diversity in shrub layer and herb layer at different elevation
2.3.2 不同海拔间α多样性指数间关系
通过对黄杉群落物种α多样性指数的相关性分析(表5)发现,灌木层和草本层的Shannon-Wiener多样性指数H与物种丰富度指数S和Simpson优势度指数D存在极显著正相关性(P<0.01),物种丰富度指数S与Simpson优势度指数D存在显著正相关性(P<0.05),Pielou均匀度指数J与其他多样性指数间相关性不显著(P<0.05)。整体来看,草本层α多样性指数间的相关性高于灌木层α多样性指数间的相关性。

表5 植物群落α多样性指数相关性分析Table 5 Correlation analysis of α diversity index of plant community
图3为相邻海拔梯度间的β多样性指数变化趋势,结果表明,随着海拔的增加,灌木层的Sørenson指数变化较大,呈双峰曲线,分别在 1943—2075 m和 2140—2185 m处达到最大值;草本层的Sørenson指数呈单峰曲线,在2140—2185 m处最大。随着海拔的上升,灌木层和草本层的Cody指数均呈单峰变化趋势,均在2075—2140 m达到最大值,但是灌木层的Cody指数变化幅度更大。上述分析表明,随着海拔梯度的增加,灌木层间物种的变化比草本层间物种的变化大。
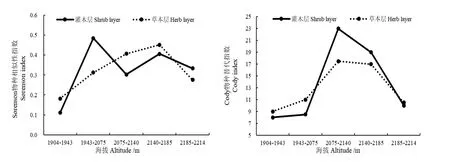
图3 相邻海拔梯度上灌木层和草本层的Sørenson指数和Cody指数Fig. 3 The Sørenson index and Cody index in shrub layer and herb layer of each two adjacent altitude gradient
由表 6可知,不同海拔梯度间灌木层物种的Sørenson指数和Cody指数有一定的差异。不同海拔梯度上,灌木层植物 Sørenson指数的平均值为0.3066,其中在海拔2075 m与海拔2214 m间最高,为 0.500;灌木层植物 Cody指数的平均值为13.2,其中在海拔2075 m和海拔2140 m间最高,为 23。但是,随着海拔差的增加,灌木层植物的Sørenson指数和Cody指数无明显变化规律。

表6 不同海拔灌木层的Sørenson指数和Cody指数Table 6 The Sørenson index and Cody index of shrub layer in different elevations
由表 7可知,不同海拔梯度间草本层物种的Sørenson指数和Cody指数也存在一定的差异。不同海拔梯度上,草本层植物Sørenson指数的平均值为0.2018,其中在海拔2140 m与海拔2185 m间最高,为0.4516;随着海拔差的增加,Sørenson指数基本上呈逐渐下降的趋势,甚至在海拔 2214 m与海拔1904 m和1943 m间以及海拔1943 m与海拔2185 m间Sørenson指数为0。不同海拔梯度上,草本层植物Cody指数的平均值为14,其中在海拔2140 m与海拔1904 m间最高,为21.5;但是,随着海拔差的增加,Cody指数无明显变化规律。
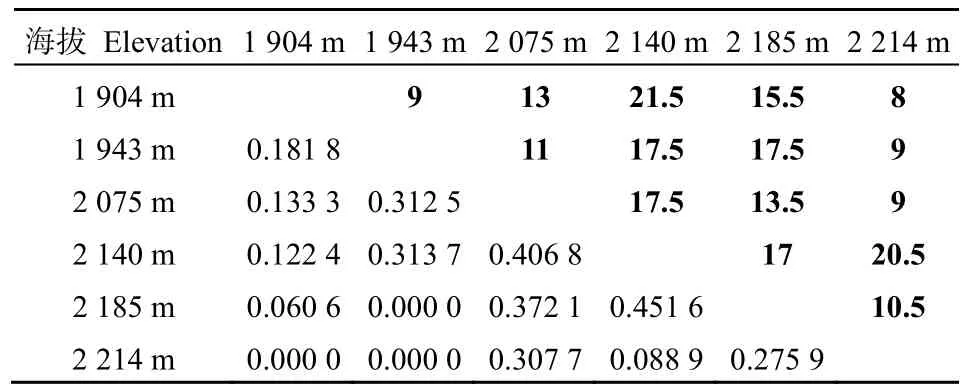
表7 不同海拔草本层的Sørenson指数和Cody指数Table 7 The Sørenson index and Cody index of herb layer in different elevations
3 讨论
3.1 黄杉群落的植物地理成分
本研究调查表明,研究区内黄杉群落植物属的地理成分复杂、多样,北温带分布区类型占明显优势(34.74%),说明黄杉群落内植物区系具有明显的温带性质,温带气候对群落内植物的系统发育过程产生了严重的影响,与研究区内的气候条件比较吻合,这与左家哺(1995)的研究结果基本一致。
3.2 海拔梯度对物种α多样性的影响
在众多生态因子中,海拔是山地环境中最为重要的因素之一,它可以通过影响太阳辐射、温度、水分和土壤等资源的分配进而影响生态系统的生态水文过程,导致植物群落空间分布格局的不同(Gaston,2000;朱珣之等,2005;张树斌等,2018)。许多研究表明,物种α多样性的海拔梯度格局主要有单调下降、单调上升、先升高后降低、先降低后升高和无规律性5种类型,负相关和中间高度膨胀型是较为普遍的格局。本研究中,黄杉群落灌木层和草本层的物种丰富度指数S、Shannon-Wiener多样性指数H、Simpson优势度指数D和Pielou均匀度指数J均呈单峰格局变化趋势,这与兰斯安等(2016)、王飞等(2018)、吴红宝等(2019)和苏闯等(2018)的研究结果相似。这可能是由于以下原因造成的:首先,相对海拔是影响喀斯特山地水热条件分布的主要因素,它通过影响光照、水分及土壤资源的再分配进而影响植物群丛的分布及结构,相对海拔较低的地带,水分充足甚至过量,但光照条件较差,生长季节光照不足常常成为限制植物分布的主要因素;相对海拔较高的地带,山体陡峭、土层较薄,气温剧烈变化产生了许多异质性的微生境,而且由于喀斯特地区特殊的双层地质结构,水分会迅速从下层流失,形成了干旱缺水的严酷生境条件,不利于植物定居;中海拔地区,水分和热量的组合、配比适宜,生境条件最好,资源的可利用率最高,优越的水热条件能使更多的物种共存,物种多样性达到最大。其次,人为干扰是另外一个原因,低海拔地区是村镇的主要分布区域,人为干扰以及牛羊等家畜的踩踏、啃食现象严重,一些物种丧失了生存机会;随着海拔的升高,人为干扰活动强度在逐渐降低,因干扰而缺失的部分物种得到自然释放,重新分布(王宇超等,2013)。
王国宏(2002)认为,群落不同生活型植物(乔、灌、草植物)的物种多样性对海拔梯度的响应不同。本研究结果表明,黄杉群落灌木层和草本层的物种α多样性指数在不同海拔上存在显著差异(P<0.05),但灌木层物种的Pielou均匀度指数不显著(P≥0.05)。这与福建梅花山常绿阔叶林植物物种多样性指数的特征相似(孔祥海等,2012)。这可能是由于环境条件(特别是水分和热量)对不同生活型植物分布的影响不同,随着海拔的变化,环境条件和资源配置发生了变化,导致了不同生活型植物的物种多样性垂直分布存在差异(Bhattarai et al.,2003;Grytnes et al.,2006)。
3.3 海拔梯度对物种β多样性的影响
本研究中,黄杉群落灌木层和草本层的物种 β多样性指数在相邻两个海拔梯度间变化很大,变化趋势不同(图3);灌木层物种Sørenson指数和Cody指数的变化幅度均比草本层大(表 6、7)。这表明研究区内灌木层物种组成在海拔梯度上差异更大,物种更替速率更快,可能与不同生活型植物的生活史策略和对环境响应不同有关(秦浩等,2019)。首先,木本植物在系统进化方面更加保守,其分布主要受历史过程的影响,表现为亲缘关系较近的物种共存;草本植物则表现为亲缘关系较远的物种共存(秦浩等,2019)。其次,木本植物受温度的影响较大(Pausas et al.,2001),木本植物的β多样性对环境差异具有相对较强的响应;草本植物受光照、水分、微地形、土壤理化性质和人为活动等影响更大(Qian et al.,2014),而且其它因子的修饰作用会削弱或加强特定生境的海拔梯度效应,草本植物的 β多样性受环境差异和地理距离的共同影响。
目前,不同学者对植物群落物种β多样性海拔梯度格局的结论不同。最普遍的格局为:随海拔高差的增加,物种β多样性指数减小。本研究表明,随着海拔高差的增加,草本层物种的Sørenson指数符合这种格局,但是,灌木层物种的Sørenson指数和 Cody指数以及草本层物种的 Cody指数均无明显变化规律。这种无明显变化规律的格局可能是由以下原因造成的:一是研究区属于喀斯特山地,黄杉群落样地的生境异质性较高,但是异质性的增加与距离的增加并不完全一致;二是世界分布种的存在可能降低了扩散限制作用(林国俊等,2010)。
综上所诉,本研究对黄杉群落多样性的海拔梯度格局进行了探讨,对保护黄杉种质资源、维持其持续更新具有重要意义。但是,研究中只对海拔梯度这一综合因子对黄杉群落物种多样性的影响进行了探讨,没有进一步深入研究水分、温度、土壤和微生物等环境因子的海拔梯度变化、对黄杉群落生物多样性的影响以及与黄杉生长发育之间的关系,以后应加强相关方面的研究。
4 结论
(1)随着海拔的升高,黄杉群落灌木层和草本层物种的α多样性指数均呈单峰格局;不同海拔上,灌木层和草本层的物种α多样性指数均存在显著差异(P<0.05),灌木层物种的Pielou均匀度指数除外。
(2)随着海拔的增加,不同生活型植物的β多样性表现出不同的变化趋势,灌木层物种组成沿海拔梯度的变化更剧烈。随着海拔高差的增加,除草本层物种的Sørenson指数逐渐减小外,其他β多样性指数变化规律不明显。
(3)相对海拔导致的水热条件差异是影响喀斯特山地物种多样性垂直分布格局的主要因素,其中,中海拔地带黄杉群落的物种多样性最高,这对黄杉种质资源的保护和维持具有重要意义。
