秦皇岛海域春季海月水母碟状幼体空间分布及其与海洋环境因子的关系
2021-09-09薛力园刘志亮宋伟安颖袁晓博陈晓
薛力园 ,刘志亮 *,宋伟,安颖 ,袁晓博 ,陈晓
1. 河北科技师范学院海洋科学研究中心,河北 秦皇岛 066004;2. 河北省海洋动力过程与资源环境重点实验室,河北 秦皇岛 066004;3. 自然资源部第一海洋研究所,山东 青岛 266061
水母是胶质浮游动物(gelatinous zooplankton)的一个重要类群,通常包括刺胞动物门(如钵水母、水螅水母、立方水母和管水母)、栉水母门的浮游类型和浮游被囊动物(Larson,1988;Mills,2001;Brotz et al.,2012)。水母身体内部的主要成分是水,碳含量小于3%,伞径大小在几微米到几米之间,生命周期较短,通常为6—9个月,从近岸浅海到深海大洋均有分布(Lucas et al.,2012;Herrmann et al.,2020)。水母是海洋生态系统的重要组成部分,它通过捕食浮游植物、浮游动物、鱼卵、仔稚鱼以及其他水母等改变或提升其营养级,成为中间甚至顶级捕食者(Purcell et al.,2001a;Boero et al.,2008;Lucas et al.,2014;Blackett et al.,2017;Lüskow,2020)。
近年来,由于全球气候变暖和人类活动(如近岸水体富营养化、过度捕捞、生物入侵和海岸工程建筑等)的影响,水母在世界不同海区大量繁殖乃至灾害性暴发的相关报道不断增加,受到了世界各国人民的普遍关注(Mills,2001;Lynam et al.,2004;Purcell et al.,2007;Uye,2008;Dong et al.,2010;Brotz et al.,2012;Condon et al.,2013;van Walraven et al.,2016;Decker et al.,2018;Yoon et al.,2019;Slater et al.,2020)。其中,钵水母是世界各地水母暴发的主要属种(Lucas et al.,2012)。水母暴发对当地生态系统和社会经济有着严重的负面影响:一方面,水母暴发会大量消耗浮游动物,摄食鱼卵和仔稚鱼,改变食物网结构,引发浮游植物大量繁殖;另一方面,水母暴发会影响商业捕鱼(如堵塞渔网、损坏渔具、倾覆船只等)、滨海养殖业(污染网箱和围栏)、旅游业(蜇伤游客)和工业基础设施(堵塞发电厂冷却水入口)等的安全运转(Møller et al.,2007;Purcell et al.,2007;Condon et al.,2011;Javidpour et al.,2016)。因此,研究水母生长变化的特征和影响因素具有重要的生态和社会经济意义(Condon et al.,2012)。
秦皇岛地处河北省东北部,南临渤海,是中国著名的海滨旅游城市,也是重要的渔业产品产出城市(张海松,2015;刘婧美等,2016)。自2008年开始,河北省沿岸海域水母数量逐年上升,秦皇岛海域暴发的水母种类主要为海月水母(Aurelia aurita)和沙海蜇(Nemopilema nomurai),暴发时间通常在 6—9 月(郑向荣等,2014;Wu et al.,2016;刘婧美等,2016)。
钵水母的生命周期大多包括底栖(螅状体)和浮游(水母体)两个阶段,导致水母种群的季节性出现和消失(Lucas et al.,2012)。生活在温带海域的钵水母成熟个体在冬季可能发生季节性萎缩或体细胞退化而消失,由存活下来的螅状体重新繁殖形成次年的水母种群(Lucas et al.,2012;Goldstein et al.,2016)。研究表明,水母幼体(如浮浪幼虫、螅状体、碟状体)的正常发育和存活是决定水母种群大小的主要因素,如一个螅状体在横裂生殖时可以产出多达40个碟状体(Schneider et al.,1994;Lucas,2001;Colin et al.,2002;Duarte et al.,2012)。探明水母种群特征及数量变动规律的一个主要环节在于掌握螅状体无性生殖和横裂生殖的控制机制以及碟状体发育成水母体的成功率(Mills,2001;Lucas et al.,2012)。螅状体横裂生殖及释放碟状体的时机、食物可利用率和非生物因素(如温度、溶解氧含量和盐度等)直接决定了碟状体的存活率(Lucas et al.,2012)。上述海洋环境因子对水母种群的影响对于理解人类活动引起的沿海生态系统的变化(如全球变化、沿岸水体富营养化和商业捕捞物种的减少等)尤为重要(Purcell et al.,1999)。
2011年,针对中国近海大型水母暴发问题启动的水母973项目对多种大型水母的生活史和海洋理化因子变化对水母不同生活史阶段的调控机制等方面进行了系统的研究(Feng et al.,2015a,2015b;Shi et al.,2015;Sun et al.,2015;Wang et al.,2015;左涛等,2016)。Sun et al.(2015)指出,水体温度和盐度的变化对水母种群数量存在显著影响。Wang et al.(2015)认为,海月水母(Aurelia sp. 1)进行横裂生殖的最适温度为 13 ℃,且食物充足时产生的碟状体营养基础好、伞径也更大。Shi et al.(2015)发现,大型水母的空间分布与小型桡足类的空间分布密切相关,其原因在于大型水母偏好摄食该浮游动物群。目前,有关秦皇岛海域水母生态学研究主要围绕水母种类组成、暴发原因和防治对策(和振武,1985;郑向荣等,2014;刘婧美等,2015;张万磊等,2015;Wu et al.,2016),而对该海域水母幼体的空间分布特征及其与环境因子的相互关系报道较少。这可能是由于水母螅状体和碟状体个体微小及其隐秘性,判定其栖息地并进行采样较为困难(Duarte et al.,2012;van Walraven et al.,2016)。
本文基于2020年4月秦皇岛海域开展的大面观测数据,分析调查海域内海月水母幼体的空间分布特征,同时分析了海月水母碟状幼体空间分布和水文环境之间的关系。本文研究结果可为进一步研究海月水母在秦皇岛海域的时空演变规律及科学制定水母防控策略提供基础数据,具有一定的科学意义和现实价值。
1 观测方法与数据
2020年4月19—23日,对秦皇岛海域(119°30′—119°55′E,39°46′—40°00′N)水母幼体和海洋水文环境进行了大面观测,共设计6条断面,每条断面5个站位,在金梦海湾内莲花岛(人工岛)增设5个加密站位,共计35个调查站位(图1)。调查期间由于站位1-1水深过浅,调查船(冀秦渔运66666,约206 kW,自质量51 t)难以到达预定位置,故该站位未进行现场采样;在a-5站位未进行现场水文环境测量,实际站位位置如图1b中黑色圆点所示。
站位水深数据来源于船载LOWRANCE HDS-7声呐探测仪,水深分布如图 1b中的实线所示。调查海域水深介于2.2—15.9 m之间,水深变化梯度总体较小。

图1 2020年4月秦皇岛海域调查站位与水深分布图Fig. 1 Investigation area with sampling locations and water depth of Qinhuangdao in April, 2020
1.1 海洋水文数据采集
调查中海洋水文数据采用一台美国 YSI多参数水质分析仪(型号YSI professional plus)测定,调查数据包括表、底层海水温度、盐度、溶解氧含量和pH。
1.2 水母幼体采集方法
水母幼体采集利用标准的浅水Ⅱ型浮游生物网,网口内径31.6 cm,网衣孔径160 μm,共取得样品 34瓶。由于水母具有昼夜垂直迁移的特性,因此采用全水柱拖网(即在各采样站位由底至表垂直拖网)来评估其种群数量。所有样品保存在 5%甲醛海水溶液中,待采样结束后返回实验室内进行镜检分析。在通风橱内将上述样品分多次转移至培养皿中,在体视镜(型号JSZ6S)下进行鉴定统计,获得水母形态、数量、伞径大小等数据。为鉴别不同生长阶段内海月水母的数量组成,本文参考Mutlu(2001)的研究结果,将海月水母划分为碟状幼体(<10 mm)和水母体(>10 mm)两个阶段。
1.3 数据分析和绘图方法
水母幼体丰度计算方法为:

式中:
ρ 为水母幼体丰度(ind·m−3);
i为属种;N为统计个体数(ind);
V为拖网水柱体积(m3)。
为了解水母幼体空间分布与海洋环境因子之间可能存在的关系,我们运用SPSS 22.0软件计算了水母幼体丰度与海水温度、盐度、溶解氧含量和pH之间的Pearson相关性系数。调查海域平均海水温度、盐度、溶解氧含量、pH和水母幼体丰度等分布图运用Surfer 13.0和Origin 2018软件绘制。
2 结果
2.1 秦皇岛海域水文环境特征
调查海域内水体温度范围在9.8—12.2 ℃之间,均值为 (11.0±0.6) ℃,最高值出现在新开河口(图2)。从平面分布上来看,调查海域内近岸水体温度高,离岸水体逐渐降低。水体盐度范围在 31.49—32.03之间,均值为 (31.84±0.14)。受河流冲淡水的影响,汤河口和新开河口水体盐度较低。溶解氧质量浓度范围在 7.70—9.56 mg·L−1之间,均值为(8.80±0.40) mg·L−1。水体溶解氧浓度在新开河口较低,在石河口、莲花岛和金山嘴外侧海域含量高。水体 pH 范围在 8.01—8.32之间,均值为(8.20±0.06)。与盐度空间变化相一致,汤河口和新开河口水体pH较低。
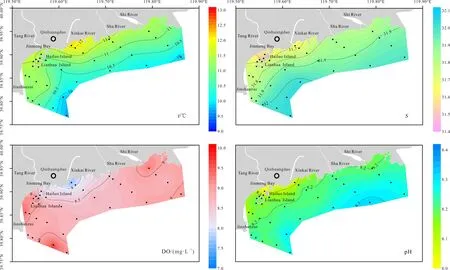
图2 2020年4月秦皇岛海域海水温度(temperature,t)、盐度(salinity,S)、溶解氧含量(dissolved oxygen,DO)和pH分布图Fig. 2 Distribution of temperature (t), salinity (S), dissolved oxygen (DO), and pH of Qinhuangdao in April, 2020
2.2 海月水母碟状幼体空间分布特征
春季调查期间,我们在调查海域内 34个站位中的11个站位发现了海月水母(Aurelia sp.),如图3所示。海月水母伞径在金梦海湾内莲花岛水域各站位最小,为1—1.5 mm;在莲花岛外侧的4-2、4-3和 5-1站位中海月水母碟状幼体伞径有所增大(2—3 mm);最大伞径(10 mm)则出现在石河口外的1-5站位(图4)。这表明调查海域内海月水母在4月尚处于碟状幼体阶段,其伞径变化呈现出以莲花岛水域为中心,向南和东北方向逐渐增大的趋势;尤其是东北侧海域伞径增大趋势更为明显。

图3 2020年4月秦皇岛海域海月水母碟状幼体照片Fig. 3 Photograph of Aurelia sp. ephyrae of Qinhuangdao in April, 2020

图4 2020年4月各调查站位海月水母碟状幼体丰度和伞径变化Fig. 4 Abundance and length distribution of Aurelia sp. ephyrae at sampling stations in April, 2020
调查海域内4月各站位海月水母碟状幼体丰度的空间分布特征如图5所示。结果表明,海月水母碟状幼体在金梦海湾内莲花岛水域丰度最大,达94 ind·m−3。而在金梦海湾以外,海月水母碟状幼体丰度迅速减小;仅在个别站位有零星分布,如 1-5、2-4和5-4站位。

图5 2020年4月秦皇岛海域海月水母碟状幼体丰度空间分布图Fig. 5 Spatial distribution of Aurelia sp. ephyrae of Qinhuangdao in April, 2020
2.3 海月水母碟状幼体空间分布与海洋环境因子的关系
由于调查海域内海月水母碟状幼体的空间分布并不均匀,我们使用lg(Y+0.001)+3对其丰度值进行标准化(Herrmann et al.,2020),其中Y为各站位水母丰度值。图6展示了调查海域内各站位海月水母碟状幼体丰度和海水温度、盐度、溶解氧含量和pH的分布信息。

图6 2020年春季秦皇岛海域各站位温度、盐度、溶解氧含量、pH和水母丰度分布Fig. 6 Variation of sea temperature (t), salinity (S), dissolved oxygen (DO), pH, and abundance of Aurelia sp. ephyrae of Qinhuangdao in spring, 2020
如图6所示,金梦海湾内莲花岛水域各站位中海月水母碟状幼体丰度高。莲花岛水域水体温度和溶解氧质量浓度均较高,分别为 (11.5±0.1) ℃和(9.12±0.10) mg·L−1;盐度较低,平均为 (31.71±0.05)。海月水母碟状幼体丰度分布中心位于莲花岛水域,这可能反映了该种水母幼体在温暖、低盐和富氧的水体环境中分布较多。
对海月水母碟状幼体与环境因子之间的相关关系进行分析,结果表明,海月水母碟状幼体丰度与水体盐度呈极显著相关性(P<0.01),与溶解氧含量和温度呈显著相关性(P<0.05),与水体pH相关性不显著(P>0.05),如图7所示。根据Pearson相关分析结果,海月水母碟状幼体丰度与盐度呈极显著负相关性(r= −0.439);海月水母碟状幼体丰度与溶解氧含量和温度呈显著正相关性,相关系数 r分别为0.369和0.348。
此外,Pearson相关分析结果显示海月水母碟状幼体伞径与水体温度之间呈极显著相关性(P<0.01),与水体盐度、溶解氧含量和 pH相关性不显著(P>0.05)。如图7所示,海月水母碟状幼体伞径与水体温度之间呈极显著负相关性,相关系数r为−0.851。
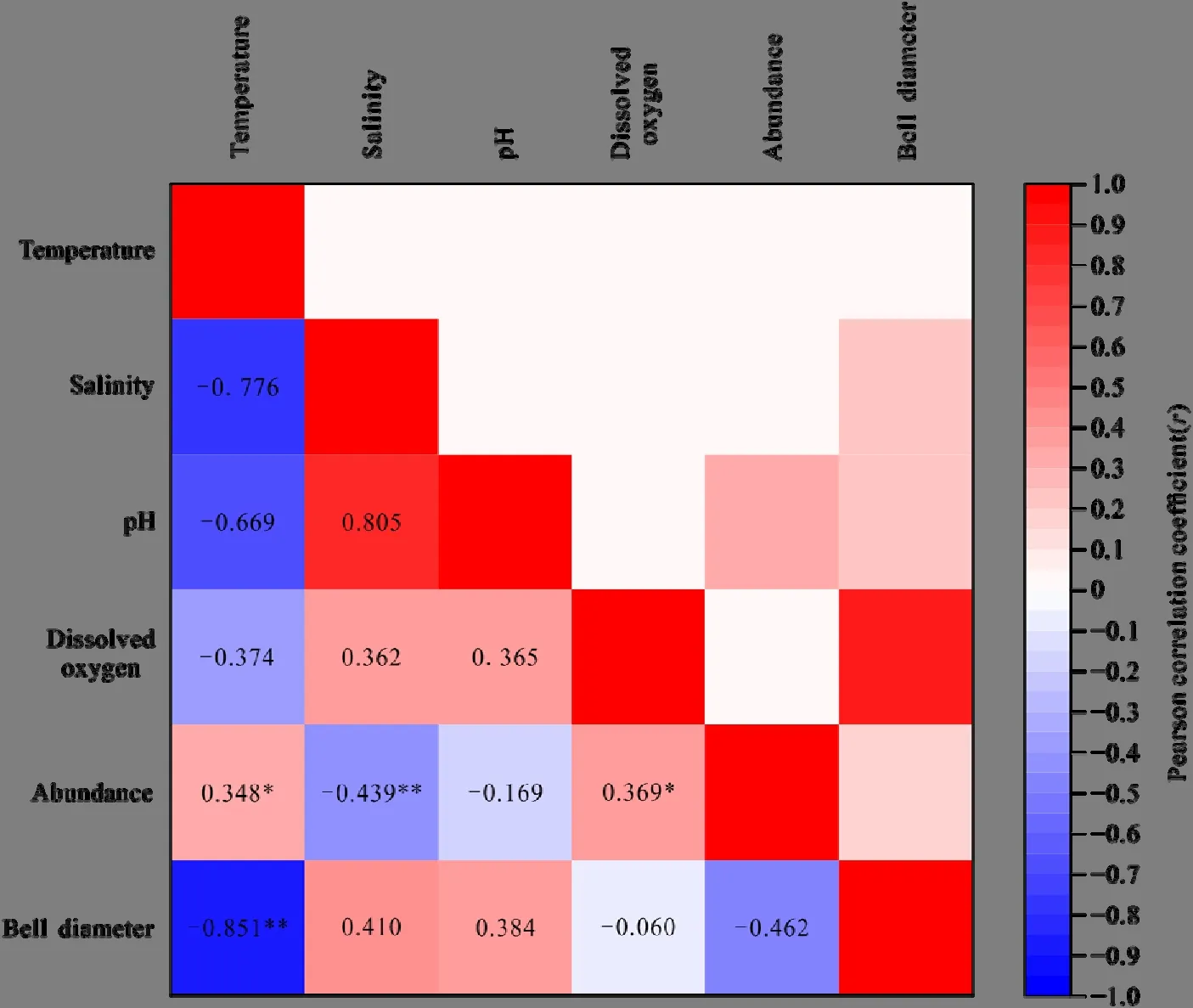
图7 2020年春季秦皇岛海域海月水母碟状幼体丰度与海洋环境因子相关性分析结果Fig. 7 Results of Pearson correlation analysis between abundance of Aurelia sp. ephyrae and sea temperature (t), salinity (S), dissolved oxygen (DO), and pH of Qinhuangdao in spring, 2020
3 讨论
水母碟状体是由环境中螅状体经横裂生殖而来,碟状体的发现可以用来指示其附近是否存在螅状体,同时水母幼体也可以用来鉴别其来源(van Walraven et al.,2016)。本文调查结果显示,秦皇岛海域4月海月水母碟状幼体分布中心位于金梦海湾内莲花岛水域。海月水母碟状幼体在莲花岛水域各站位丰度最高,伞径最小。而在金梦海湾以外,海月水母碟状幼体丰度较金梦海湾水域明显减少,且伞径较大。金梦海湾内莲花岛水域的海月水母碟状幼体可能来源于本地水母螅状体种群,其空间分布可能与碟状体的集中释放有关。此外,莲花岛水域的海月水母碟状幼体可以通过海流输送到离岸海域;在此过程中,海月水母碟状幼体丰度迅速减小,伞径逐渐增大(Han et al.,2019)。秦皇岛海域近岸水体温度高,离岸海域逐渐降低,因此海月水母碟状幼体伞径与温度之间呈负相关关系。
海洋环境因子(如温度、盐度和光照等)的变化能够影响水母螅状体的无性生殖过程,在控制水母种群数量和分布中起着极为重要的作用(Graham et al.,2001;Purcell,2007)。调查海域内春季(4月)水体温度、盐度和溶解氧含量等环境因子的空间分布存在明显差异。海月水母碟状幼体分布中心位于金梦海湾内莲花岛水域,表明该海域相对温暖、低盐和富氧的水体环境有利于海月水母幼体的生长、存活和繁殖。
相关性分析结果显示,海月水母碟状幼体丰度与水体盐度相关性最强,其次为溶解氧含量和温度,与海水pH相关性不显著。盐度是影响海月水母无性生殖过程的环境因子之一(Lucas et al.,2012)。Purcell et al.(2007)指出,海月水母(Aurelia labiata)螅状体发生横裂生殖的数量随盐度降低(34—27)而增加;产生碟状体的数量也随之增加。Holst et al.(2010)发现,海月水母(Aurelia aurita)螅状体发生横裂生殖的最适盐度为28,此时产生碟状体的数量也最多;盐度升高或降低均会产生不利影响。但是,一些学者认为盐度对海月水母(Aurelia aurita)的繁殖和生长速率并无显著影响(Watanabe et al.,2001;Willcox et al.,2007)。但 Willcox et al.(2008)认为,盐度可能与环境中营养盐输入和食物可利用率有关,而食物可利用率与钵水母的出芽生殖速率存在正相关关系。Purcell et al.(2007)也指出,盐度变化可以通过影响海洋生产力进而影响水母种群的数量。金梦海湾内水体严重富营养化,而这与营养物质的增加、营养物质比率的改变密切相关,也往往导致了该海域浮游生物量的增加(Purcell et al.,2007;Han et al.,2019)。一般来说,海月水母(Aurelia aurita)种群数量与浮游动物生物量的季节变化密切相关,其种群大小取决于食物可利用率(Goldstein et al.,2016)。Decker et al.(2018)指出,白令海东部水域浮游动物生物量在时间序列上的峰值先于水母生物量的增加,表明食物可利用率是导致该海域水母种群波动的关键因素。在丹麦Kertinge Nor水域,小型水母的大量繁殖造成的食物短缺导致了海月水母最大伞径仅为几厘米,而在其他海域通常为30 cm(Riisgård et al.,2010)。调查海域内 4月盐度整体变化不大,受河流冲淡水影响,位于汤河口附近的莲花岛水域水体盐度较低 (31.71±0.05),但处于海月水母幼体生长的适宜范围内。综上所述,调查海域内水体盐度对海月水母碟状幼体的空间分布存在显著影响,莲花岛水域低盐的水体环境为海月水母碟状幼体的大量聚集提供了有利条件。
除盐度外,调查海域内海月水母碟状幼体丰度与水体溶解氧含量也呈显著相关性。Keister et al.(2000)认为底层海水溶解氧含量能够影响浮游动物在整个水柱中的丰度和深度分布,浮游动物在低氧水体(<2 mg·L−1)中数量稀少,而在溶解氧含量大于2 mg·L−1的水体环境中数量较多。研究表明,大多数水母属种生活在溶解氧含量大于2 mg·L−1的水体环境中,但一些属种(如 Chrysaora quinquecirrha和Mnemiopsis leidyi)对低氧水体表现出耐受性(Purcell et al.,2001b)。Ishii et al.(2008)发现,海月水母螅状体在低氧水体(<2.86 mg·L−1)能够存活并产生新的螅状体,当溶解氧含量由2.86 mg·L−1上升到 6.43 mg·L−1后,螅状体开始横裂生殖并释放碟状体,但释放的碟状体数量无明显差异。一般来说,低氧水体会对浮游动物生物量和丰度产生不利影响(Keister et al.,2020)。Condon et al.(2001)指出,Chrysaora quinquecirrha螅状体进行无性生殖的比例和产生碟状体的数量随溶解氧含量降低而减少。秦皇岛海域春季(4月)水体溶解氧含量整体较高(>2 mg·L−1),有利于海月水母幼体的存活、生长和繁殖。据此,本文认为莲花岛水域富氧的水体环境也是海月水母碟状幼体大量聚集的有利条件。
此外,相关性分析结果还显示了海月水母碟状幼体丰度与水体温度呈显著相关性。研究表明,海水温度与水母种群的分布和生物量之间存在正相关关系,随着全球变暖,海水温度升高,暖温性水母种群的分布范围将会扩大,生物量也将相应增加(Purcell,2005;Willcox et al.,2008;Holst,2012;曲长凤等,2014)。在温带海域,海月水母(Aurelia aurita)螅状体通常在冬末春初温度和日照量开始升高时进行横裂生殖(Lucas,2001;Purcell,2007;Liu et al.,2009)。万艾勇等(2012)发现,胶州湾内的海月水母碟状体通常在4月(水温为11.4 ℃)开始出现,在 6月(水温为 20.6 ℃)碟状体丰度达到最高,主要集中出现在红岛附近水域。而在日本东京湾内,海月水母碟状体主要出现在冬季 12月至次年 3月,在春季和夏初发育成水母体(Watanabe et al.,2001)。在实验室条件下,海月水母螅状体在 10—14 ℃时进行横裂生殖,其个体和种群的生长速率随温度升高而显著增加(Willcox et al.,2007;Treible et al.,2019)。莲花岛水域水体温度平均为 (11.5±0.1) ℃,有利于海月水母螅状体进行横裂生殖。Purcell(2007)发现,生活在北太平洋的海月水母(Aurelia labiata)螅状体在10 ℃时进行横裂生殖的数量比在7 ℃时多4倍,且温度每升高1 ℃,释放碟状体的数量增加11.3%。Liu et al.(2009)也认为,海月水母(Aurelia aurita)螅状体产生碟状体的数量随温度升高而增加,如25 ℃和30 ℃时产生碟状体的数量比20 ℃时多一倍。室内受控实验和野外调查结果均表明,食物充足和温暖的水体环境有利于水母螅状体的横裂生殖过程,并产生更多的碟状体(Purcell,2007;Liu et al.,2009;Lucas et al.,2012)。此外,温度还能显著影响海月水母碟状幼体的生长速率,如东北太平洋海域的海月水母(Aurelia labiata)碟状体的最适生长温度范围为 12—21 ℃,而黄海北部的海月水母碟状体最适生长温度为25 ℃(Widmer,2005;付志璐等,2011)。大量研究表明,温度是影响海月水母螅状体横裂生殖和水母幼体生长速率的关键环境因子之一,并在一定时空尺度上影响水母的丰度和分布(Lucas,2001;Lynam et al.,2004;Purcell et al.,2007;Treible et al.,2019)。本研究海月水母碟状幼体丰度与温度的相关性最低,可能是由于秦皇岛海域春季(4月)水体温度变化不大,处于海月水母螅状体横裂生殖和碟状体生长的适宜温度范围内。
4 结论
通过对秦皇岛海域2020年4月航次调查结果的总结分析,我们初步了解了该海域春季海月水母碟状幼体数量、伞径大小、空间分布及其与环境因子的关系,获得主要结论如下:
(1)调查期间,秦皇岛海域内海月水母主要以碟状体形式存在,伞径介于 1—10 mm。金梦海湾莲花岛水域海月水母碟状幼体伞径最小(1—1.5 mm),丰度最高(94 ind·m−3)。在金梦海湾以外,海月水母碟状幼体伞径逐渐增大,丰度呈迅速减小趋势。海月水母碟状幼体分布中心位于金梦海湾内莲花岛水域。上述特征表明,秦皇岛海域的海月水母起源于金梦海湾内的本地螅状体种群。
(2)秦皇岛海域内海月水母碟状幼体丰度与海水盐度呈极显著负相关性(P<0.01),与温度和溶解氧含量呈显著相关性(P<0.05),与海水pH相关性不显著。相关性分析结果表明,海月水母碟状幼体丰度的空间分布与盐度相关性最强,其次为溶解氧含量和温度。金梦海湾内莲花岛水域相对温暖、低盐和富氧的水体环境有利于海月水母螅状体进行横裂生殖,为海月水母碟状幼体的存活和生长提供了良好的栖息场所。
致谢:感谢“冀秦渔运 66666”号全体船员为现场样品的采集所付出的艰辛努力。
