海岛棉新海35花药发育及花粉受精的研究
2021-06-19杨静马益赞王为然闵玲宁新民孔杰
杨静,马益赞,王为然,闵玲,宁新民,孔杰*
(1.华中农业大学作物遗传改良国家重点实验室,武汉430070;2.新疆农业科学院经济作物研究所,乌鲁木齐830091)
作为雌雄同株的植物,生长环境、成花数量、花芽发育等都会影响棉花纤维品质。花药的正常发育作为花粉粒成熟及受精的必备条件,同样影响棉纤维产量和品质。对花药发育的研究需从花器官发育过程的细胞结构[1]及其具体的细胞特征进行探究。自1999年,Sanderszai等[2]以拟南芥为研究对象,对花药发育过程进行了详细的时期划分后,人们研究其他开花植物花发育过程时,均以拟南芥花药内部细胞组成特征作为参照来确定花药发育时期。虽然花药发育不同时期,环境条件的改变能够决定花药发育早期细胞的命运,但花药细胞的特殊结构对花器官的发育及部分功能的行使具有识别作用[1,3-4]。不同的开花时间、各个花器官对环境条件的敏感程度[5]、植物本身的部分氧化物及相关基因都会影响开花植物花芽的正常发育[6],因此,棉花花芽发育时期、花蕾大小及其花器官的细胞发育特征成为研究棉花花发育的关键。其中花药的发育过程不仅包括多种细胞形态建成,也是后期双受精完成的先决条件。近年,部分学者研究了陆地棉的生育期[7]、花期[8]和高温胁迫对花药发育的影响[9],而针对花药发育过程中细胞结构、相关发育时期[10]及受精过程的研究并不多见。其中海岛棉花蕾发育与花药细胞的形态建成、发育时期以及花粉的双受精过程尚未报道。本研究利用半薄切片技术与显微镜观察方法,确定新海35花蕾发育过程中各发育时期的细胞特征,建立花发育时期与花蕾大小的对应关系,同时对花粉萌发及受精过程进行了探究,以期为海岛棉花发育时期的判定及相关细胞发育的研究提供参考。
1 材料及方法
1.1 试验材料
试验材料为新海35,由新疆农业科学院经济作物研究所提供。2017—2018年分别种植于新疆维吾尔自治区巴音郭楞蒙古自治州库尔勒市阿瓦提乡新疆农业科学院经济作物研究所育种家基地(86.08°E,41.68°N)和华中农业大学作物遗传改良国家重点实验室的棉花温室。库尔勒大田白天最低气温为16℃,最高气温37℃;温室最低气温20℃,最高气温36℃,温室采用花盆单株种植,所用土壤为砾石与基质1∶2混合,常规水肥、防虫管理。田间管理同常规大田,定期施肥浇水。根据长期的监测观察,新海35号从播种到开花为61~67 d,总生育期为135~140 d。
1.2 试验方法
1.2.1花药取样。在花芽发育早期进行挂牌标记,以便后期对应相同生育期取样。2017年7月2日和2018年6月15日,分别取大田材料和温室材料制作切片。从花芽出现开始,待花芽发育3~6 d后,根据花蕾发育长度,每增加1 mm取样1次,5个重复。取回的花蕾去掉苞叶,用游标卡尺从底端蜜腺处至花蕾顶部进行长度测量、记录、拍照,并进行解剖观察。
1.2.2雌蕊柱头取样。2018年7月4-8日,在库尔勒大田每日16:00对发育正常且长势一致的新海35号去雄,次日10:00选取即将开放且发育一致的花蕾对前1 d去雄后留存的雌蕊柱头进行授粉,随后套上纸袋,避免其他花粉的污染。人工授粉后0、5、10、20、30、45 m in及1、1.5、2、4、8、10、12、14、16、18、20、22、24、26、28、30 h的雌蕊柱头取样;从授粉后30 h开始,之后每隔4 h对授粉的柱头取1次,即34、38、42 h……74 h。且每次每个时间点采集5个重复,冰盒保湿带回实验室用于后续切片的固定。
1.2.3半薄切片制作、染色及观察。样品用FPA固定液(丙酸∶福尔马林∶70%乙醇=5∶5∶90,体积比)固定;依次用30%、50%、70%、80%乙醇脱水(体积比)分别脱水15 m in,最后用无水乙醇漂洗1次。用预渗透液(无水乙醇与渗透液1∶1(体积比)混合,1 g的HarderⅠ溶于100 m L Technovit 7100制成渗透液)预渗透12 h,再用渗透液渗透24 h,然后使用包埋液(渗透液∶HarderⅡ=15∶1,体积比)进行包埋。
切片、展片、染色:将完成后熟的树脂块进行切片,厚度5μm。将切好的半薄切片放置在涂有多聚赖氨酸的载玻片上,吸去多余的水,将切好的花药切片用0.5%甲苯胺蓝染色。
封片、拍照:染色完成后,蒸馏水清洗3次,晾干水渍用中性树胶封片,待树胶完全干燥后用ZEISSAxioscope A1显微镜进行拍照观察。
2 结果与分析
2.1 海岛棉花蕾与花药发育形态变化
海岛棉的开花周期较长,从花芽出现至开花一般需要28~30 d。新海35在花芽发育3~5 d后,花蕾长度为2~3 mm(图1B)。当花芽发育6~8 d时,花蕾长度为4~6 mm,此时萼片的发育早于花瓣,且完全包裹花瓣。花芽发育14~18 d时,花蕾长度为6~13 mm,花瓣被萼片半包裹。当花芽发育24~26 d,花蕾长度为14~21 mm,此时被萼片包裹着的花瓣发育速度明显加快,且超过了萼片(图1B)。随着花蕾继续生长,在花芽发育26~27 d,花蕾长度为21~22mm,此时萼片的发育速度较为缓慢,花瓣继续快速生长体积变大,花瓣颜色由最初的灰白色(1~12 mm)变为灰绿色(12~20 mm),而后为淡黄色,快开花时变为柠檬黄色(图1B)。花药颜色从最初的白色(<12 mm)逐渐变为金黄色(图1B)。花丝和花柱随着花药的发育逐渐伸长,在开花前1天花瓣、花丝、柱头和花药生长迅速。在开花当天,花药开裂,花丝快速伸长(图1C-E),伸长的花丝在花药开裂后将成熟的花粉粒散落至柱头(图1E),花粉开始萌发,花粉管伸长,并进行双受精过程。开花后1~2 d花柱不再伸长,此时花粉受精过程完成(图1F),卵细胞“休眠”。合子时期,花瓣则变为橘红色(图1F),花药开始失水(图1G),至此海岛棉花蕾内部花器官发育完成。

图1 新海35花蕾及雄蕊发育形态Fig.1 The development of buds and stamen of G.barbadense Xinhai35
2.2 花蕾大小与花药的发育时期的关系
2.2.1花原基与雄蕊原基时期。棉花的花芽分化需要经过花原基分化、苞片分化、花萼分化、花瓣分化、雄蕊分化、雌蕊分化、药隔形成、花粉母细胞形成、减数分裂、花粉粒形成等过程。当花芽出现2~3 d可以观察到长2~3 mm的三角形苞片;花蕾发育至0.5 mm左右时,苞片生长较快,且苞片迅速增大,其生长速度明显快于萼片。当萼片在显微镜下明显可见时,萼片上部边缘形成的突起即花瓣(图2A),此时花瓣的顶部生长较底部迅速,且在其上部的左右两端有可见的突起(图2A)。当花芽出现3~4 d,花蕾长度0.5~1 mm时,雄蕊原基已经出现(图2B)。此时萼片生长速度比花瓣快,且萼片体积增大后包裹着其它正在发育的花器官。随着发育的进程,花瓣向左右相互交错覆盖,包裹着隆起的分生组织,而分生组织内缘上部形成的突起即雄蕊原基(图2B)。随着花芽发育,雄蕊上形成的突起逐渐弯向中央(图2B)。雄蕊继续向上生长,中央部分分化出雌蕊原基(图2C)。
2.2.2花药孢原细胞与花药四分体时期。新海35花芽出现3~5 d,花蕾长度为2~3 mm,此时经由雄蕊原基中间层即L2层细胞分化的孢原细胞已清晰可见(图2D)。待花芽出现4~6 d,花蕾长至3~4 mm时,孢原细胞经平周分裂形成周缘细胞与造孢细胞(图2E)。花芽出现5~7 d,花蕾长至4~5 mm,此时次生周缘细胞分化出花药的中间层和绒毡层,内皮层出现,裂口组织与维管组织形成,裂口区域和连接区清晰可见(图2F)。花芽出现7~9 d花蕾长至5~6 mm时,经花药原基分化的花粉母细胞和4层花药壁形成,花粉母细胞结构清晰,4层花药壁从外到内依次为:表皮层、药室内壁、中间层和绒毡层。此时花药被药隔分隔为4个药室,花药形态呈清晰的蝴蝶状(图2G)。当花芽出现10~12 d时,花蕾生长至6~7 mm,花粉母细胞开始进行减数分裂(图2H),花药中含有贮藏物质的一层细胞开始减少,随后细胞逐渐趋于扁平,直至发生解体。花芽出现12~13 d花蕾生长至7~8 mm时,花粉母细胞进入减数分裂后期。随着花粉母细胞的逐渐成熟,可见被胼胝质包裹、含有单倍体小孢子的四分体形成(图2I)。当花芽出现13~14 d,花蕾生长至8~9 mm时,胼胝质降解,小孢子从花粉母细胞的四分体中释放出来,形成成熟的小孢子,至此小孢子的整个发育阶段完成(图2J)。
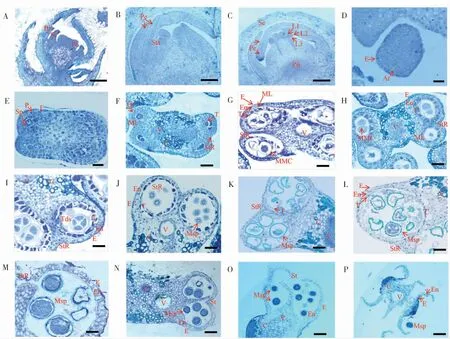
图2 新海35花蕾长度与花药发育时期的对应关系Fig.2 The relationship between buds length and anther development stages of Xinhai35
图2 (续)Fig.2 Continued
2.2.3花药绒毡层发育时期。绒毡层作为种子植物花药幼时最内侧的细胞层,能够为小孢子的发育与外壁形成提供蛋白质和碳水化合物等营养及原料,合成并分泌胼胝体酶、识别蛋白和孢粉素。当新海35的花芽出现5~7 d,花蕾发育至4~5 mm时,次生周缘细胞分化出绒毡层(图2F),花芽出现7~9 d,花蕾为5~6 mm时,绒毡层完全形成(图2G)。在花芽出现12~13 d,花蕾为7~8 mm时,绒毡层细胞出现液泡化,中层细胞受到挤压,细胞质浓度增加,细胞核体积增大,胼胝质积累,中层细胞开始降解(图2I)。在花芽出现15~17 d,花蕾长度为8~9 mm时,绒毡层合成并分泌胼胝体酶,溶解胼胝质使小孢子从四分体中释放。花芽发育18~22 d,花蕾9~11 mm时,来源于绒毡层的孢粉素填充在小孢子的初生外壁,小孢子体积增大,花药膨胀生长。花蕾发育24~25 d,花蕾14~24 mm时,孢粉素积累,使花药壁增厚,单核花粉粒经非均等有丝分裂形成双核花粉粒(图2M-N),此时绒毡层完全降解。
2.2.4花粉粒的形成与释放。海岛棉花芽发育18~22 d,花蕾长度为9~11 mm时,从四分体释放出来的小孢子只有1个细胞核,同时具有大液泡。花芽出现22~24 d,花蕾为11~14 mm时,绒毡层逐渐降解从而为小孢子提供营养,同时部分蛋白在小孢子外壁积累(图2L),形成花粉壁的前体。当花芽发育24~25 d,花蕾生长至14~24 mm时,小孢子进行有丝分裂,花药中有丰富的内含物,花粉粒开始膨胀生长,可以清晰地观察花粉粒结构(图2M)。当花蕾长度大于24 mm时,花粉粒逐渐成熟,随后裂口组织开始降解,待花粉粒完全成熟,花药开裂,花粉粒沿裂口区域冲出药室,开始散粉(图2N-O)。
2.3 花粉萌发与受精
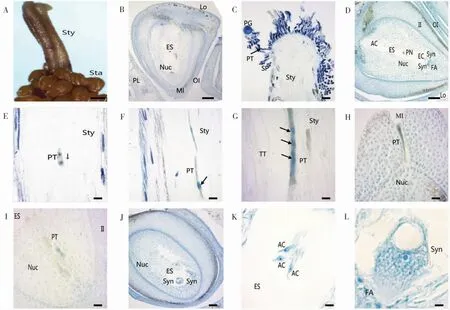
图3 新海35花粉萌发与花粉管伸长过程Fig.3 Pollen germ ination and pollen tube elongation of Xinhai 35
2.3.1花粉粒萌发与花粉管伸长。海岛棉花药正常开裂后,花粉散出,通过人工授粉将花粉涂抹于去雄的柱头,一定时间后取柱头,制作切片进行观察。经过0.5~1 h,花粉粒开始萌发,可清晰地观察到花粉管伸出(图3 C),并沿花柱的引导组织逐渐向下呈不规则的曲线形生长(图3E-G)。大约在授粉后8 h,可以清晰看到花粉管中携带的生殖细胞发生分裂,形成前后排列的2个子细胞(图3E)。授粉后12~14 h,花粉管伸长至花柱末端,并进入子房(图3H)。授粉后16~20 h,花粉管已伸长到达胚珠的珠孔,并通过珠孔进入珠心(图3H)。授粉后20~24 h,花粉管进一步到达胚囊处,并释放出营养细胞和生殖细胞等内容物(图3 I-J)。
2.3.2双受精过程。当人工授粉24 h后,显微镜观察胚囊中清晰可见1枚精子正在与卵细胞进行受精(图4A);另1个精子则先靠近1个极核并与之结合(图4B),随后与另1极核融合形成受精极核(图4C-E),至此双受精完成。双受精后可以在胚囊内清晰看见1个合子和1个助细胞(图4E)。
2.3.3胚乳发育。海岛棉的柱头授粉后50 h,观察到含2个游离核的胚乳(图4D);授粉后74 h,可看见已经形成了较多的游离核分布在胚囊内(图4F)。与此同时,受精卵结束休眠,胚乳进行第1次分裂。其主要特点是:基细胞大于顶细胞,基细胞靠近珠孔,细胞相对较大;而顶细胞远离珠孔,细胞相对比较小(图4G)。此时纤维伸长也比较明显(图4H)。
3 讨论
3.1 棉花生长周期与花蕾形态
花器官的形态建成在植物发育过程中扮演着重要的作用,其表型不仅指示环境条件的改变,而且关系植株的生殖发育。本研究中海岛棉新海35从播种到开花需要61~67 d,从现蕾到开花需要28~32 d,从播种到吐絮的总生育期为135~140 d。而陆地棉从现蕾到开花一般仅需要25~30 d[11]。从播种到吐絮的生长周期也比海岛棉明显短4~8 d[7],有研究认为,这与种质相关基因及其棉酚含量有关[12]。新海35花蕾、萼片、花瓣和雄蕊在开花前5 d快速生长,与陆地棉的研究结果相似[13]。随发育进程,新海35的花蕾、萼片、花瓣和雄蕊、花药颜色随之改变(图1B-D),且花瓣和花药颜色变化比陆地棉更为明显[14]。这可能与次生代谢过程中花青素、类黄酮和花色素等相关物质的积累有关[15]。海岛棉生殖发育过程中,光合作用产生的干物质以及氮素等营养物质均用于生殖生长前期,导致后期积累较少,不能满足快速发育需要,所以生育期较陆地棉长[16]。除此之外,海岛棉对光照时间、光照强度及温度有着严苛的要求,管理措施不当会导致开花与结铃时期延后[17]。因此,海岛棉新海35的生长周期、花蕾和花药颜色、发育表型变化是对自然环境条件的响应,也是种质资源、不同地理来源和遗传变异共同作用所产生的结果[8,14]。
3.2 棉花花蕾大小与花药发育时期
棉花现蕾后,花药的发育需经历雄蕊原基突起、孢原细胞分化、1次平周分裂形成造孢细胞,再经有丝分裂形成花粉母细胞。花粉母细胞进行减数分裂形成小孢子,小孢子形成成熟花粉粒[18-19]。海岛棉H276保持系花蕾长度3~4 mm时,为花粉母细胞时期;花蕾4~5 mm时为四分体时期;花蕾长度5~6 mm为小孢子单核早期,绒毡层开始退化;花蕾6~7 mm时绒毡层完全降解,部分小孢子已经发育成为成熟的花粉粒。花蕾>7 mm时形成成熟花粉粒[20]。而陆地棉四分体时期花蕾长度为4.1~5.5 mm,单核时期花蕾纵径5.6~6.0 mm,成熟的花粉粒时期花蕾纵径为8.1~10.0 mm[21]。其它研究发现,在棉花开花期,造孢细胞时期花蕾<3.5 mm,花粉母细胞时期花蕾为3.5~5.0 mm,花粉粒时期花蕾>5 mm[22]。本研究中新海35花蕾长度至5~6 mm、7~8 mm、8~9 mm和11~14 mm分别对应花药发育中的花粉母细胞形成期、四分体时期、小孢子释放期和绒毡层降解时期,均比相同发育时期陆地棉的花蕾长。可能是为海岛棉生殖发育后期积累营养物质的要求[16]。
3.3 棉花花粉粒的发育
花粉粒的成熟是雄蕊发育的关键,只有花粉粒正常发育,才能保证植株的受精和结实。在雄蕊原基中初生造孢细胞分化形成花粉母细胞[23]。次生周缘细胞分化形成包裹着花粉母细胞的三层同心圆花药壁[24],再经对称分裂形成中间层与绒毡层。当小孢子形成时,绒毡层细胞为其提供所需营养物质,并将一些遗传物质传递给小孢子[25]。小孢子的发育会直接影响花粉粒的成熟。棉花蕾期花粉粒的发育受影响会导致花粉败育。研究证实,陆地棉花蕾处于开花前15~16 d为四分体时期至小孢子发育早期,开花前9~11 d为单核晚期至双核早期的发育阶段[26]。而新海35花芽出现10~13 d,花蕾生长至6~7 mm和7~8 mm时分别为花粉母细胞减数分裂和四分体时期,该时期花蕾的花药细胞最易受非生物胁迫影响。本研究发现,海岛棉新海35在花蕾为8~9 mm时,小孢子从四分体胼胝质中释放出来。当花蕾11~14 mm时,绒毡层开始降解,为小孢子的发育提供营养物质,而细胞残余及脂类物质等覆盖在小孢子表面形成花粉包被[27]。陆地棉花蕾9~14 mm时,合成并分泌孢粉素,为花粉发育提供营养物质[28],而海岛棉新海35则在花蕾11~14 mm完成这一过程。新海35绒毡层降解起始时的花蕾比陆地棉1355B稍大[10],可能是由于温度、光照及种质资源的相关调控基因改变了绒毡层细胞程序性死亡的时间[29]。因此,海岛棉花蕾长度5~6 mm和7~8 mm对应花粉母细胞和四分体时期的正常发育,在花蕾8~9 mm和11~14 mm时绒毡层能否进行正常降解关系着花粉粒成熟和花药的正常开裂[30]。
3.4 棉花花粉萌发及受精过程
在陆地棉中双受精是精卵融合以及与极核的融合。其中,极核融合的机制是:外膜在多个点上的直接融合,即内质网的融合;随后是内核膜的融合形成桥,随后桥扩大并愈合,逆向释放被网罗的细胞质成分,最后形成新的完整的核膜,完成核的融合[16]。精卵细胞的融合与极核的融合方式基本一致[25]。本研究中,海岛棉花粉粒在柱头上经过0.5~1 h后开始萌发,花粉管开始生长。授粉后8 h,可以清晰看到花粉管中携带的生殖细胞发生分裂。授粉后20~24 h,花粉管到达胚囊。授粉后24 h,精子与卵细胞进行受精。授粉50 h后,形成游离胚。74 h,胚乳进行第一次分裂,此时纤维明显伸长。有研究发现,授粉后12 h和24 h观察到的花粉管数量基本一致,且30℃最适于花粉管萌发,花柱中花粉管的平均数量最多[31]。研究发现,花粉萌发率可作为响应温度的主要指标[32]。在30℃培养条件下花粉萌发率最高,25℃培养条件下的花粉管长度最长。不同温度条件不同材料的花粉萌发率和花粉管长度不同。同时花粉萌发率和花粉管长度也反映了单株铃数和成铃率[33]。有研究发现,授粉后15 h花粉管通过珠孔进入胚囊,释放内含物。48 h后可见胚的分裂。72 h后可以明显观察到分裂的早期胚[34]。但在新海35中授粉50 h后,才观察到纤维发育,这可能是因为海岛棉的受精过程中,极核的受精是精子先与1个极核结合,然后再与另1个极核进行核膜、核质的融合,因此所需要受精时间较长。这也证实了海岛棉的受精时间比陆地棉长[25]。双受精完成之后,棉花纤维进入伸长期,与此同时纤维细胞出现大液泡[35]。这一时期是遗传物质传递的关键时期,环境条件及相关基因的作用都会影响海岛棉受精时间。
4 结论
海岛棉新海35花药颜色和花药细胞随着花蕾发育呈现动态变化。在相同的花药细胞发育阶段,新海35的花蕾比陆地棉的花蕾更长,且各发育阶段所需的时间更长。结合花芽发育时间、花蕾形态学及细胞学观察,确定了新海35花粉母细胞形成期、四分体时期、小孢子释放期和绒毡层降解时期花蕾长度分别为5~6 mm、7~8 mm、8~9 mm和11~14 mm阶段。同时新海35授粉后,花粉粒萌发需要0.5~1 h,花粉管延伸到子房需要12~14 h,而受精过程则是在授粉后24 h完成,而授粉后50~74 h胚乳开始发育并伴随着纤维的发育。
